


******************************
Pleurotomaria
| Pleurotomaria Temporal range: Jurassic–Cretaceous | |
|---|---|
 | |
| Pleurotomaria niloticiformes fossil at the Geological Museum in Copenhagen | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Mollusca |
| Class: | Gastropoda |
| (unranked): | clade Vetigastropoda |
| Superfamily: | Pleurotomarioidea |
| Family: | Pleurotomariidae |
| Genus: | Pleurotomaria Sowerby, 1821 [1] |
Pleurotomaria is a genus of sea snails, marine gastropod mollusks in the family Pleurotomariidae.[2]
Species
Species within the genus Pleurotomaria include
- Species brought into synonymy
- † Pleurotomaria adansoniana Crosse & P. Fischer, 1861: synonym of Entemnotrochus adansonianus (Crosse & P. Fischer, 1861)
- † Pleurotomaria africana Tomlin, 1948: synonym of Bayerotrochus africanus (Tomlin, 1948)
- † Pleurotomaria beyrichii Hilgendorf, 1877: synonym of Mikadotrochus beyrichii (Hilgendorf, 1877)
- † Pleurotomaria hirasei Pilsbry, 1903: synonym of Mikadotrochus hirasei (Pilsbry, 1903)
- † Pleurotomaria quoyana P. Fischer & Bernardi, 1856: synonym of Perotrochus quoyanus quoyanus (P. Fischer & Bernardi, 1856)
- † Pleurotomaria rumphii Schepman, 1879: synonym of Entemnotrochus rumphii (Schepman, 1879)
- † Pleurotomaria salmiana Rolle, 1899: synonym of Mikadotrochus salmianus (Rolle, 1899)
- † Pleurotomaria westralis Whitehead, 1987: synonym of Bayerotrochus westralis (Whitehead, 1987)
With current taxonomic changes the genus Pleurotomaria has been reserved exclusively for fossil species (denoted with † below).
Known fossil species of Pleurotomaria include:[3]
- † Pleurotomaria agarista Billings 1865
- † Pleurotomaria anglica Sowerby 1818 (synonym: Trochus anglicus)
- † Pleurotomaria angulosa d'Orbigny 1842
- † Pleurotomaria antitorquata Münster 1840
- † Pleurotomaria arctica Toula 1875
- † Pleurotomaria arenaria Girty 1908
- † Pleurotomaria awakinoensis Begg and Grant-Mackie 2003
- † Pleurotomaria barrealensis Cowper Reed 1927 (synonym: Neoplatyteichum barrealensis)
- † Pleurotomaria bicoronata Sandberger and Sandberger 1855
- † Pleurotomaria biondii Gemmellaro 1889
- † Pleurotomaria bodana Roemer 1855
- † Pleurotomaria brennensis Reed 1932
- † Pleurotomaria calcifera Billings 1859
- † Pleurotomaria cancellata Stauffer 1909
- † Pleurotomaria carinifera Girty 1908
- † Pleurotomaria catherinae Gemmellaro 1889
- † Pleurotomaria cingulata Goldfuss 1844
- † Pleurotomaria coheni Gemmellaro 1889
- † Pleurotomaria comata Lindström 1884
- † Pleurotomaria coniformis de Koninck 1883
- † Pleurotomaria costulatocanaliculata Sandberger and Sandberger 1853
- † Peurotomaria discoidea Girty 1908
- † Pleurotomaria doris Hall 1862
- † Pleurotomaria elderi Girty 1908
- † Pleurotomaria estella Hall and Whitfield 1872
- † Pleurotomaria exaltata d’Archiac and de Verneuil 1842
- † Pleurotomaria gracilis Phillips 1841
- † Pleurotomaria halliana Shumard 1859
- † Pleurotomaria hectori Trechmann 1918 (synonym: Pleurotomaria (Sisenna) hectori)
- † Pleurotomaria helicoides Cowper Reed 1901
- † Pleurotomaria hisingeri Goldfuss 1864
- † Pleurotomaria hyale Billings 1865
- † Pleurotomaria isaacsii Hall and Whitfield 1873
- † Pleurotomaria isomorpha Gemmellaro 1889
- † Pleurotomaria karetai Begg and Grant-Mackie 2003
- † Pleurotomaria kiritehereensis Begg and Grant-Mackie 2003
- † Pleurotomaria mariani Gemmellaro 1889
- † Pleurotomaria mazarensis Gemmellaro 1889
- † Pleurotomaria mica Girty 1908
- † Pleurotomaria monilifera Terquem and Jourdy 1873
- † Pleurotomaria murchisoniaeformis Gemmellaro 1889
- † Pleurotomaria neglecta Girty 1908
- † Pleurotomaria neosolodurina Dacque 1905
- † Pleurotomaria nikitini Fliegel 1901
- † Pleurotomaria notlingi Koken 1896
- † Pleurotomaria nuda Delpey 1941
- † Pleurotomaria obliqua Fliegel 1901
- † Pleurotomaria otapiriensis Begg and Grant-Mackie 2003
- † Pleurotomaria perfasciata Hall 1860
- † Pleurotomaria pericarinata Hall 1847 (synonyms: Cyclonema pericarinata, Gyronema pericarinatum)
- † Pleurotomaria perlata Hall 1852
- † Pleurotomaria perornata Shumard 1859
- † Pleurotomaria perversa Whidborne 1889
- † Pleurotomaria planulata Girty 1908
- † Pleurotomaria plicifera d'Eichwald 1860
- † Pleurotomaria postumia Billings 1865
- † Pleurotomaria proutiana Shumard 1859
- † Pleurotomaria psiche Gemmellaro 1889
- † Pleurotomaria putilla Girty 1908
- † Pleurotomaria replicata Lindström 1884
- † Pleurotomaria retroplicata Gemmellaro 1889
- † Pleurotomaria richardsoni Girty 1908
- † Pleurotomaria roemeri Koken 1889 (synonym: Euryzone roemeri)
- † Pleurotomaria salomonensis Gemmellaro 1889
- † Pleurotomaria sigaretus Sandberger and Sandberger 1855
- † Pleurotomaria striatissima Cowper Reed 1901
- † Pleurotomaria subsulcata Goldfuss 1844
- † Pleurotomaria subtilistriata Hall 1847
- † Pleurotomaria trinchesii Gemmellaro 1889
- † Pleurotomaria tunstallensis King 1848
- † Pleurotomaria viola Billings 1865
- † Pleurotomaria virgo Billings 1865
- † Pleurotomaria waimumu Begg and Grant-Mackie 2003
- † Pleurotomaria wurmi Römer, 1843
- † Pleurotomaria zitteli Holzapfel 1882
References
- Jump up ^ J. Sowerby, Mineral Conchology iii, p. 139, t. 278, Dec. 1821.
- Jump up ^ Pleurotomaria Sowerby, 1821. Retrieved through: World Register of Marine Species on 3 April 2011.
- Jump up ^ Paleobiology Database: Pleurotomaria
- Schmidt, W. & O. Bellec (1994). Findings of some uncommon sea-shells off Madagascar. African Journal of Tropical Hydrobiology and Fisheries 5(1): 63 - 66
- J. G. Begg and J. A. Grant-Mackie. 2003. New Zealand and New Caledonian Triassic Pleurotomariidae (Gastropoda, Mollusca). Journal of the Royal Society of New Zealand 33:223-268
Gastropod shell



The gastropod shell is a shell which is part of the body of a gastropod or snail, one kind of mollusc. The gastropod shell is an external skeleton or exoskeleton, which serves not only for muscle attachment, but also for protection from predators and from mechanical damage. In land snails, in some freshwater snails and in intertidal marine snails, the shell is also an essential protection against the sun, and against drying out.
Most gastropod shells are spirally coiled. The coiling is usually right-handed, but in some taxa the coiling is left-handed and in a very few species there can be both right-handed and left-handed individuals.
The gastropod shell has several layers, and is typically made of calcium carbonate precipitated out into an organic matrix known as conchiolin. The shell is secreted by a part of the molluscan body known as the mantle.
Some gastropods are shell-less (slugs), but the majority of gastropods do have a shell. In almost every case the shell consists of one piece, and is typically spirally coiled, although some groups, such as the various families and genera of limpets, have simple cone-shaped shells as adults.
The study of mollusc shells, including gastropod shells, is called conchology.
Contents
[hide]Chirality in gastropods
Because coiled shells are asymmetrical, they possess a quality called chirality, the "handedness" of an asymmetrical structure.
By far the majority (over 90%)[1] of gastropod species have dextral (right-handed) shells in their coiling, but a small minority of species and genera are virtually always sinistral (left-handed), and a very few species (for example Amphidromus perversus[2]) show an even mixture of dextral and sinistral individuals.
In species that are almost always dextral, very rarely a sinistral specimen will be produced, and these oddities are avidly sought after by some shell collectors.
If a coiled gastropod shell is held with the spire pointing upwards and the aperture more or less facing the observer, a dextral shell will have the aperture on the right-hand side, and a sinistral shell will have the aperture on the left-hand side.
This chirality of gastropods is often overlooked when photographs of coiled gastropods are "flipped" by a non-expert prior to being used in a publication. This image "flipping" results in a normal dextral gastropod appearing to be a rare and abnormal sinistral one.
The chirality in gastropods appears in early cleavage (spiral cleavage) and the gene NODAL is involved.[3] A more recent study (2013) correlates the asymmetric coiling of the shell by the left-right asymmetric expression of the decapentaplegic gene in the mantle.[4]
Mixed coiling populations
In few cases, both left- and right-handed coiling are found in the same population.[5] Sinistral mutants of normally dextral species and dextral mutants of normally sinistral species are rare but well documented occurrences among land snails in general.[5] Populations or species with normally mixed coiling are much rarer, and, so far as is known, are confined, with one exception, to a few genera of arboreal tropical snails.[5] Besides Amphidromus, the Cuban Liguus vittatus (Swainson), Haitian Liguus virgineus (Linnaeus) (family Orthalicidae), some Hawaiian Partulina and many Hawaiian Achatinella (family Achatinellidae), as well as several species of Pacific Island Partula (family Partulidae), are known to have mixed dextral-sinistral populations.[5] The independent appearance of this variation in unrelated groups is probably the result of a simple mutation, whose primary import is with physiological adaptations to arboreal life and not with the direction of coiling.[5] In Partula both dextral and sinistral embryos have been recovered from the same brood pouch, although normally all embryos coil in the same direction.[5] In Amphidromus there is no information on the heredity of this character.[5]
A possible exception may concern some of the European clausiliids of the subfamily Alopiinae.[5] They are obligatory calciphiles living in isolated colonies on limestone outcrops.[5] Several sets of species differ only in the direction of coiling, but the evidence is inconclusive as to whether left- and right-handed shells live together.[5] Soos (1928, pp. 372–385)[full citation needed] summarized previous discussions of the problem and concluded that the right- and left-handed populations were distinct species.[5] Others have stated that these populations were not distinct, and the question is far from settled.[5] The Peruvian clausiliid, Nenia callistoglypta Pilsbry (1949, pp. 216–217),[full citation needed] also has been described as being an amphidromine species.[5]
The genetics of reverse coiling in a rare dextral mutant of another clausiliid, Alinda biplicata (Montagu), has been studied by Degner (1952).[5] The mechanism is the same as in Radix peregra (Müller), with the direction of coiling determined by a simple Mendelian recessive.[5] Any change in direction caused by cross-fertilization is delayed one generation by an unknown mechanism.[5]
Formation of the shell
Morphology
 | | |
| Morphology of typical spirally coiled shell. The shell of Zonitoides nitidus, a land snail, has dextral coiling. Upper image: Dorsal view, showing whorls and apex | Photo of the shell of Zonitoides nitidus with an apical view, an apertural view and a basal view |
Gastropod shell morphology is usually quite constant among individuals of a species, and with exceptions, fairly constant among species within each family of gastropoda. Controlling variables are:
- The rate of growth per revolution around the coiling axis. High rates give wide-mouthed forms such as the abalone, low rates give highly coiled forms such as Turritella or some of the Planorbidae.
- The shape of the generating curve, roughly equivalent to the shape of the aperture. It may be round, for instance in the turban shell, elongate as in the cone shell or have an irregular shape with a siphonal canal extension, as in the Murex.
- The rate of translation of the generating curve along the axis of coiling, controlling how high-spired the resulting shell becomes. This may range from zero, a flat planispiral shell, to nearly the diameter of the aperture.
- Irregularities or "sculpturing" such as ribs, spines, knobs, and varices made by the snail regularly changing the shape of the generating curve during the course of growth, for instance in the many species of Murex.
- Ontologic growth changes as the animal reaches adulthood. Good examples are the flaring lip of the adult conch and the inward-coiled lip of the cowry.
Some of these factors can be modelled mathematically and programs exist to generate extremely realistic images. Early work by David Raup on the analog computer also revealed many possible combinations that were never adapted by any actual gastropod.
Some shell shapes are found more often in certain environments, though there are many exceptions. Wave-washed high-energy environments, such as the rocky intertidal zone, are usually inhabited by snails whose shells have a wide aperture, a relatively low surface area, and a high growth rate per revolution. High-spired and highly sculptured forms become more common in quiet water environments. The shell of burrowing forms, such as the olive and Terebra, are smooth, elongated, and lack elaborate sculpture, in order to decrease resistance when moving through sand.
A few gastropods, for instance the Vermetidae, cement the shell to, and grow along, solid surfaces such as rocks, or other shells.
Standard ways of viewing a shell

Apertural view of shell of Valvata sincera

Abapertural view of shell of Valvata sincera

Basal or umbilical view of shell of Valvata sincera

This dorsal view of the living animal Calliostoma bairdii also shows the dorsal view of its shell




In photographs or illustrations, a gastropod shell can be shown oriented in a number of standard ways:
- apertural view: this is the most common viewing angle. The shell is shown in its full length with its aperture facing the viewer and the apex at the top. When the aperture is on the right side, then the shell is called "right-handed" or dextral; if the aperture is on the left side, the shell is called "left-handed" or sinistral.
- abapertural view: the shell is shown in its full length with its aperture 180° away from the viewer, and with the apex at the top.
- apical view (or dorsal view): the shell is shown looking down directly onto the apex
- basal view (or umbilical view): the shell is shown viewed directly from the base. In most cases where there is an umbilicus, this is in clear view.
Description
The shell begins with the minute embryonic whorls of the protoconch, which is often quite distinct from the rest of the shell. From the protoconch, which forms the apex of the spire, the coils or whorls of the shell gradually increase in size. Normally the whorls are circular or elliptical in section, but from compression and other causes a variety of forms can result. The spire can be high or low, broad or slender according to the way the coils of the shell are arranged, and the apical angle of the shell varies accordingly. The whorls sometimes rest loosely upon one another (as in Epitonium scalare). They also can overlap the earlier whorls such that they may be largely or wholly covered by the later ones. When an angulation occurs, the space between it and the suture above it constitutes the area known as the "shoulder" of the shell. The shoulder angle may be simple or keeled, and may sometimes have nodes or spines.
The most primitive sculpture of the gastropod shell consists of revolving ridges or spirals, and of transverse folds or ribs. Primary spirals appear in regular succession on either side of the first primary, which generally becomes the shoulder angle if angulation occurs. Secondary spirals appear by intercalation between the primary ones, and generally are absent in the young shell, except in some highly accelerated types. Tertiary spirals are intercalated between the preceding groups in more specialized species. Ribs are regular transverse foldings of the shell, which generally extend from the suture to suture. They are usually spaced uniformly and crossed by the spirals. In specialized types, when a shoulder angle is formed, they become concentrated as nodes upon this angle, disappearing from the shoulder above and the body below. Spines may replace the nodes in later stages. They form as notches in the margin of the shell and are subsequently abandoned, often remaining open in front. Irregular spines may also arise on various parts of the surface of the shell (see Platyceras). When a row of spines is formed at the edge or outer lip of the shell this sometimes remains behind as a varix as in (Murex) and many of the Ranellidae. Varices may also be formed by simple expansion of the outer lip, and a subsequent resumption of growth from the base of the expansion. These simple varices may project from the shell (Epitonium) or be reflected backwards (Harpa). Periodic enlargements of ribs are not considered as varices.
The aperture or peristome of the shell may be simple or variously modified. An outer and an inner (columellar) lip are generally recognized. These may be continuous with each other, or may be divided below by an anterior notch. This, in some types (Fusinus, etc.) it is drawn out into an anterior siphonal canal, of greater or lesser length.
An upper or posterior notch is present in certain taxa, and this may result in the formation of a ridge or shelf next to the suture (Clavilithes). An outer (lateral) emargination or notch, sometimes prolonged into a slit occurs in certain types (Pleurotomidae, Pleurotomaridae, Bellerophontidae, etc.), and the progressive closing of this slit may give rise to a definitely marked slit band. In some cases the slit is abandoned and left as a hole (Fissurellidae), or by periodic renewal as a succession of holes (Haliotis). The outer emargination is often only indicated by the reflected course of the lines of growth on the shell.
On the inside of the outer lip, various ridges or plications called lirae are sometimes found, and these occasionally may be strong and tooth-like (Nerinea). Similar ridges or columellar plicae or folds are more often found on the inner lip, next to the columella or central spiral twist. These may be oblique or normal to the axis of coiling (horizontal), few or numerous, readily seen, or far within the shell so as to be invisible except in broken shells. When the axis of coiling is hollow (perforate spire) the opening at the base constitutes the umbilicus. The umbilicus varies greatly in size, and may be wholly or in part covered by an expansion or callus of the inner lip (Natica).
Many Recent shells, when the animal is alive or the shell is freshly empty, have an uppermost shell layer of horny, smooth, or hairy epidermis or periostracum, a proteinaceous layer which sometimes is thick enough to hide the color markings of the surface of the shell. The periostracum, as well as the coloration, is only rarely preserved in fossil shells.
The apertural end of the gastropod shell is the anterior end, nearest to the head of the animal; the apex of the spire is often the posterior end or at least is the dorsal side. Most authors figure the shells with the apex of the spire uppermost. In life, when the soft parts of these snail are retracted, in some groups the aperture of the shell is closed by using a horny or calcareous operculum, a door-like structure which is secreted by, and attached to, the upper surface of the posterior part of the foot. The operculum is of very variable form in the different groups of snails that possess one.[6]
Parts of the shell

The terminology used to describe the shells of gastropods includes:
- Aperture: the opening of the shell
- Lip = peristome: the margin of the aperture
- Apex: the smallest few whorls of the shell
- Body whorl (or last whorl): the largest whorl in which the main part of the visceral mass of the mollusk is found
- Columella: the "little column" at the axis of revolution of the shell
- Operculum: the "trapdoor" of the shell
- Parietal callus: a ridge on the inner lip of the aperture in certain gastropods
- Periostracum: a thin layer of organic "skin" which forms the outer layer of the shell of many species
- Peristome: the part of the shell that is right around the aperture
- Plait: folds on the columella.
- Protoconch: the nuclear whorls; the larval shell, often remains in position even on an adult shell
- Sculpture: ornamentation on the outer surface of a shell
- Lira: one kind of shell sculpture
- Siphonal canal: an extension of the aperture in certain gastropods
- Spire: the part of the shell above the body whorl.
- Suture: The junction between whorls of most gastropods
- Teleoconch : the entire shell without the protoconch; the postnuclear whorls.
- Umbilicus: in shells where the whorls move apart as they grow, on the underside of the shell there is a deep depression reaching up towards the spire; this is the umbilicus
- Varix: on some mollusk shells, spaced raised and thickened vertical ribs mark the end of a period of rapid growth; these are varices
- Whorl: each one of the complete rotations of the shell spiral
Shape of the shell
The distinction of the shape of the shell may vary, based on the purpose. For example distinguishing into three groups can be based on the height - width ratio:[7]
- oblong - the height is much bigger than the width
- globose or conical shell - the height and the width of the shell are approximately the same
- depressed - the width is much bigger than the height
The following are the principal modifications of form in the gastropod shell.[8]
- Regularly spiral:
- Bulloid : bubble-shaped Bulla ampula
- Coeloconoid : a slightly concave conical shell in which the incremental angle increases steadily during growth (see: Calliostoma)
- Cone-shaped, obconic. Conus
- Contabulate, short, with shouldered whorls
- Convolute ; aperture as long as the shell, nearly or quite concealing the spire. Cypraea
- Cylindrical, pupiform. Lioplax, Pupa
- Cyrtoconoid: approaching a conical shape but with convex sides (see: Gibbula)
- Depressed, lenticular. Ethalia carneolata
- Discoidal. Elachorbis
- Ear-shaped. Haliotis
- Elongated, subulate, elevated. Terebra
- Few-whorled. Helix pomatia.
- Fusiform, spindle-shaped. Fusinus
- Gibbous. Whorls swelled beyond the normal contour of increase (usually on the aperture side ). Streptaxis.
- Globular. Natica
- Many-whorled. Millerelix peregrina.
- Short, bucciniform. Buccinum
- Trochiform, pyramidal, conical with a flat base. Trochus
- Turbinated ; conical, with rounded base. Turbo
- Turrited, turriculated, babylonic ; an elongated shell with the whorls angulated or shouldered on their upper part. Turritella
- Scalariform, whorls not impinging. Epitonium
- Irregularly spiral, evolute. Siliquaria, Vermetus
- Tubular. Dentalium, or tooth-shell.
- Shield-shaped. Umbraculum
- Boat-shaped, slipper-shaped. Crepidula
- Conical or limpet-shaped. Patella
- Biconic : two conical shapes touching their bases and tapering at both ends : Fasciolaria tulipa
- Multivalve and imbricated. Chiton

oblong shell of Bulgarica denticulata

globose shell of Sphincterochila candidissima

depressed shell of Escargot de Quimper


Detailed distinction of the shape can be:[9][10]

cap shape

ear shape

neritiform

planispiral

depressed trochiform or valvatiform

trochiform

ovate-conic

conic

elongate-conic or turriform or cockscrew shape

top shape

spindle shape - the sea snail Syrinx aruanus has the largest shell of any living gastropod.

club shape

barrel shape

egg shape

irregular shape














Pear shape means both shapes: ovate-conic and conic.

Dimensions
The most frequently used measurements of a gastropod shell are: the height of the shell, the width of the shell, the height of the aperture and the width of the aperture. The number of whorls is also often used.
In this context, the height (or the length) of a shell is its maximum measurement along the central axis. The width (or breadth, or diameter) is the maximum measurement of the shell at right angles to the central axis. Both terms are only related to the description of the shell and not to the orientation of the shell on the living animal.
The largest height of any shell is found in the marine snail species Syrinx aruanus, which can be up to 91 cm.[11]
The central axis is an imaginary axis along the length of a shell, around which, in a coiled shell, the whorls spiral. The central axis passes through the columella, the central pillar of the shell.
Evolutionary changes
Among proposed roles invoked the variability of shells during evolution include mechanical stability,[12] defense against predators[13] (evolution of shells against external attacks has been proved by nanoindentation),[14] sexual selection[15] and climatic selection.[16][17]
The shell of some gastropods have been reduced or partly reduced during the evolution. This reduction can be seen in all slugs, in semi-slugs and in various other marine and non-marine gastropods. Sometimes the reduction of the shell is associated with predatory way of feeding.
Some taxa even lost the coiling of their shell during evolution.[18] According to Dollo's law, it is not possible to regain the coiling of the shell after it is lost. Despite that, there are few genera in the family Calyptraeidae that changed their developmental timing (heterochrony) and gained back (re-evolution) a coiled shell from the previous condition of an uncoiled limpet-like shell.[18]
Taphonomic implications
In large enough quantities, gastropod shells can have enough of an impact on environmental conditions to affect the ability of organic remains in the local environment to fossilize.[19] For example, in the Dinosaur Park Formation, fossil hadrosaur eggshell is rare.[19] This is because the breakdown of tannins from local coniferous vegetation would have caused the ancient waters to become acidic.[19] Eggshell fragments are present in only two microfossil sites, both of which are predominated by the preserved shells of invertebrate life, including gastropods.[19] It was the slow dissolution of these shells releasing calcium carbonate into the water that raised the water's pH high enough to prevent the eggshell fragments from dissolving before they could be fossilized.[19]
Variety of forms

Turritella communis, many-whorled shell of tower snail

X-ray image of Turritella

Shell of marine cowry snail - Cypraea nebrites

X-ray image of Cypraea




X-ray image of the shell of Tonna galea
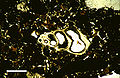
Thin section in plane-polarized light of microscopic gastropod shell, from Holocene lagoonal sediment of Rice Bay, San Salvador Island, Bahamas. Scale bar 500 µm.




References
This article incorporates public domain text from references [5][6] and CC-BY-2.0 text from reference.[17]
- Jump up ^ Schilthuizen M. & Davison A. (2005). "The convoluted evolution of snail chirality". Naturwissenschaften 92(11): 504–515. doi:10.1007/s00114-005-0045-2.
- Jump up ^ Amphidromus perversus (Linnaeus, 1758)
- Jump up ^ Myers P. Z. (13 April 2009) "Snails have nodal!". The Panda's Thumb, accessed 3 May 2009.
- Jump up ^ Keisuke Shimizu, Minoru Iijima, Davin HE SetiamargaIsao Sarashina, Tetsuhiro Kudoh, Takahiro Asami, Edi Gittenberger, Kazuyoshi Endo; Left-right asymmetric expression of dpp in the mantle of gastropods correlates with asymmetric shell coiling, EvoDevo 2013, 4:15
- ^ Jump up to: a b c d e f g h i j k l m n o p q
 Laidlaw F. F. & Solem A. (1961). "The land snail genus Amphidromus: a synoptic catalogue". Fieldiana Zoology 41(4): 505-720.
Laidlaw F. F. & Solem A. (1961). "The land snail genus Amphidromus: a synoptic catalogue". Fieldiana Zoology 41(4): 505-720. - ^ Jump up to: a b Grabau A. W. & Shimer H. W. (1909) North American Index Fossils Invertebrates. Volume I.. A. G. Seiler & Company, New York. pages page 582-584.
- Jump up ^ Falkner G., Obrdlík P., Castella E. & Speight M. C. D. (2001). Shelled Gastropoda of Western Europe. München: Friedrich-Held-Gesellschaft, 267 pp.
- Jump up ^ George Washington Tryon, Structural and systematic conchology, 1882, p. 43-44
- Jump up ^ Hershler R. & Ponder W. F.(1998). "A Review of Morphological Characters of Hydrobioid Snails". Smithsonian Contributions to Zoology 600: 1-55. [1].
- Jump up ^ Dance P. S. (). Shells.
- Jump up ^ Wells F. E., Walker D. I. & Jones D. S. (eds.) (2003) "Food of giants – field observations on the diet of Syrinx aruanus (Linnaeus, 1758) (Turbinellidae) the largest living gastropod". The Marine Flora and Fauna of Dampier, Western Australia. Western Australian Museum, Perth.
- Jump up ^ Britton J. C (1995) "The relationship between position on shore and shell ornamentation in 2 size-dependent morphotypes of Littorina striata, with an estimate of evaporative water-loss in these morphotypes and in Melarhaphe neritoides". Hydrobiologia 309: 129-142. abstract.
- Jump up ^ Wilson A. B., Glaubrecht M. & Meyer A. (March 2004) "Ancient lakes as evolutionary reservoirs: evidence from the thalassoid gastropods of Lake Tanganyika". Proceedings of Royal Society London Series B - Biological Sciences 271: 529-536. doi:10.1098/rspb.2003.2624.
- Jump up ^ Bignardi C., Petraroli M. & Pugno N. M. (2010). "Nanoindentations on Conch Shells of Gastropoda and Bivalvia Molluscs Reveal Anisotropic Evolution Against External Attacks". Journal of Nanoscience and Nanotechnology 10(10): 6453-6460. doi:10.1166/jnn.2010.2626.
- Jump up ^ Schilthuizen M. (5 June 2003) "Sexual selection on land snail shell ornamentation: a hypothesis that may explain shell diversity". BMC Evolutionary Biology 3: 13. doi:10.1186/1471-2148-3-13.
- Jump up ^ Goodfriend G. A. (1986) "Variation in land-snail shell form and size and its causes – a Review". Systematic Zoology 35: 204-223.
- ^ Jump up to: a b Pfenninger M., Hrabáková M., Steinke D. & Dèpraz A. (4 November 2005) "Why do snails have hairs? A Bayesian inference of character evolution". BMC Evolutionary Biology 5: 59. doi:10.1186/1471-2148-5-59
- ^ Jump up to: a b Collin R. & Cipriani R. (22 December 2003) "Dollo’s law and the re-evolution of shell coiling". Proceedings of the Royal Society B 270(1533): 2551-2555. doi:10.1098/rspb.2003.2517 PMID 14728776.
- ^ Jump up to: a b c d e Tanke, D.H. and Brett-Surman, M.K. 2001. Evidence of Hatchling and Nestling-Size Hadrosaurs (Reptilia:Ornithischia) from Dinosaur Provincial Park (Dinosaur Park Formation: Campanian), Alberta, Canada. pp. 206-218. In: Mesozoic Vertebrate Life—New Research Inspired by the Paleontology of Philip J. Currie. Edited by D.H. Tanke and K. Carpenter. Indiana University Press: Bloomington. xviii + 577 pp.
Schneckenhaus


Als Schneckenhaus oder -gehäuse wird die spiralförmig gewundene kalkige Schale der Schnecken bezeichnet. Es ist mit zwei Muskeln mit dem Körper der Schnecke verbunden und dient dem Schutz des Weichtierkörpers vor Fressfeinden und Verletzungen.
Inhaltsverzeichnis
Evolution der Schneckengehäuse
Das Schneckengehäuse entstand im Verlaufe eines langen, bereits im Kambrium einsetzenden evolutionären Prozesses aus Kalkstacheln und Schalenplatten früher Weichtiere. Die sich daraus gebildete Schale ist allen Angehörigen der Conchifera eigen. Im Unterschied zu den Schalen der Muscheln (Bivalvia), Kahnfüßer (Scaphopoda) und Kopffüßer (Cephalopoda) ist die Schale der Schnecken, bei aller Verschiedenheit in der Gestalt und mancherlei äußeren Ähnlichkeiten zu Schalen dieser Weichtiergruppen, jedoch stets spiralig gewunden.
Bau und Form der Schale
Die Bezeichnung „Gewinde“ umfasst das Protoconch und das Teleoconch bis einschließlich des vorletzten Umgangs. Der letzte Umgang mit der Mündung wird als Basis bezeichnet.
Protoconch

Die Schale entsteht im Embryonalstadium der Schnecke. Der in dieser Entwicklungsstufe angelegte Abschnitt der Schale bildet die Spitze (Apex) des Gehäuses, von dessen weiteren Verlauf es sich deutlich unterscheiden kann. Die Gestalt des Embryonalgewindes oder Protoconchs ist ein Merkmal, das bei der Artbestimmung (insbesondere in der Paläontologie, da so gut wie nie Weichteile erhalten sind) eine bedeutsame Rolle spielt. Dieser Teil des Gehäuses ist meist glatt, kann aber auch skulptiert sein.
Das Protoconch kann sein:
- orthostroph (auch homöostroph) = das Embryonalgewinde hat den gleichen Windungssinn wie das Teleoconch (der häufigste Fall)
- heterostroph = mit entgegengesetztem Windungssinn (weitaus seltener)
- alliostroph = Das Embryonalgewinde verläuft im Verhältnis zum Teleoconch versetzt in einem Winkel von bis 90° (in diesem Fall „liegt“ es quasi auf dem übrigen Gewinde)
- paucispiral = mit 1 bis 2 Umgängen
- multispiral = mit drei oder mehr Umgängen
Teleoconch
Der auf das Protoconch folgende Abschnitt, der den weitaus größten Teil des Gehäuses einnimmt, wird als Teleoconch bezeichnet. Die Anwachsstreifen und andere die Skulptur prägende Merkmale des Teleoconchs treten meist artspezifisch in sehr variantenreichen Formen auf. Siehe hierzu die Erläuterungen in den betreffenden Abschnitten weiter unten.
Innerer Aufbau der Schale

Die Schneckenschale ist mehrschichtig aufgebaut. Von innen nach außen tragen die Schichten die Bezeichnungen: Hypostracum, Ostracum und Periostracum. Die beiden erstgenannten Schichten bestehen aus Aragonit, einem Kalkmaterial (CaCO3). Sie bilden das stabile „Grundgerüst“ der Schale. Das Periostracum hingegen, das auch als Schalenhaut bezeichnet werden kann, besteht aus einem komplexen Protein, dem so genannten Conchiolin (auch Conchin genannt). Diese organische äußere Schutzschicht der Schale kann bei adulten Schnecken nicht erneuert werden, da die an der Mündung sitzenden Drüsen, in denen das Conchiolin produziert wird, nur während des Wachstums der Schneckenschale arbeiten. Gleichwohl können Schnecken Verletzungen ihrer Schale von innen heraus mit Hilfe Kalk abscheidender Drüsenzellen reparieren, die „Reparaturstelle“ weist jedoch aufgrund des Fehlens des Periostracums eine zumeist deutlich sichtbare gröbere Oberflächenstruktur auf.
Wenn an der Innenseite der Schale dünne Aragonit-Plättchen abgelegt sind, kann diese innere Schalenschicht (das Hypostracum) durch Lichtbrechung schillernde Effekte hervorrufen. Dieser als Perlmutt bekannte Teil der Schale tritt bei einigen Süß- und Salzwasserschnecken auf, insbesondere bei den Seeohren (Haliotis).
Die Spiralform der Schale

Die Schneckenschale verläuft vom Apex bis Mündung um die eigene Achse, die als Spindel (Columella) bezeichnet wird, wenn die Umgänge sich berühren. Anderenfalls entsteht ein trichterförmiger Hohlraum, der Nabel (Umbilicus). Der Grund für die Entstehung dieser spiralförmigen Schalengestalt ist in der Ontogenese der Schnecken zu suchen: Eine Seite des Eingeweidesacks wächst schneller als die andere, wodurch eine Drehbewegung auftritt, die wiederum die Spiralform und die Windungsrichtung der Schale determiniert. Dieser Vorgang wird als Torsion bezeichnet.
Die Schale der Schnecken kann – zumeist aber nicht immer artspezifisch – rechtsgewunden (dextral), wie bei den weitaus meisten Schneckenarten, oder linksgewunden (sinistral) sein. Wenn beim aufrecht (mit dem Apex nach oben) stehenden Gehäuse die dem Betrachter zugewandte Mündung links liegt, heißt es linksgewunden, im umgekehrten Falle rechtsgewunden. Linksgewundene Weinbergschnecken werden im Volksmund als Schneckenkönige bezeichnet.
Weitere Schalenmerkmale
Größe, Form und Musterung der Schale sind oft artspezifisch und mithin bei der Bestimmung von Bedeutung.
Größe
Aquatisch lebende Schnecken bringen infolge des Auftriebs im Wasser tendenziell größere Formen hervor als Landbewohner. Insbesondere unter den Meeresschnecken kommen sehr große Arten mit zum Teil sehr schweren Gehäusen vor, wie beispielsweise die im Meeresgebiet zwischen Indonesien und Australien vorkommende bis 1 m Gehäusehöhe erreichende Große Rüsselschnecke (Syrinx aruanus). Andererseits sind unter den rezenten Schnecken Formen anzutreffen, deren Gehäusegröße deutlich unterhalb eines Millimeters liegt (z. B. die Angehörigen der Familie Omalogyridae).
Skulptierung

Die Oberfläche der Schale weist eine mehr oder minder deutliche Skulptur auf, deren Merkmale sehr unterschiedlich stark ausgeprägt sein können.
Sie besteht im einfachsten Fall aus Anwachsstreifen, die in der Wachstumsphase entstehen. Der Verlauf dieser Anwachsstreifen auf dem Gehäuse wird wie folgt beschrieben (Sichtweise jeweils auf das aufrecht stehende, also mit dem Apex nach oben zeigende Gehäuse mit Blick auf die Mündung – wie bei der nebenstehend abgebildeten Semicassis granulata):
- orthoclin = geradlinig von oben nach unten,
- prosoklin = leicht bogenförmig von links oben nach rechts unten,
- opistoklin = leicht bogenförmig von oben rechts nach unten links,
- prosocyrt = bumerangförmig mit der Ausbuchtung nach links,
- opistocyrt = bumerangförmig mit der Ausbuchtung nach rechts.
Ein anatomisches Merkmal, das mit den Anwachsstreifen eng zusammenhängt, sind die so genannten Varices (Sing.: Varix). Dabei handelt es sich um verdickte axiale Rippen, die gewöhnlich in gleichmäßigen Intervallen auf den Umgängen auftreten und als eine Verdickung der Außenlippe während eines Ruhezustands im Wachstum der Schnecke ausgebildet worden sind. Daran ist erkennbar, dass das Wachstum der Schnecke in Schüben stattgefunden hat (siehe nebenstehende Abbildung).
Neben Anwachsstreifen und Varices können Ritzen, Furchen, Streifen, Kiele, Rippen, Knoten, Stacheln und Wülste auf der Schalenoberfläche auftreten. Verlaufen solche Merkmale parallel zur Sutur werden sie als Spiralverzierung bezeichnet. Sehr feine Linien oder Furchen werden Lirae (Sing.: Lira) genannt. Sofern die Schalenverzierungen schräg oder senkrecht (also rechtwinklig zur Sutur) angeordnet sind, spricht man von Querverzierung.
Die Mündung[Bearbeiten]

Die Mündung bildet den Abschluss des letzten Umgangs (Körperumgang, Basis). Der Einfachheit halber wird im Folgenden der Mündungsrand in drei Bereiche gegliedert:
- Die Parietalis (Mündungswand): Dieser Bereich liegt unmittelbar unter dem vorletzten Umlauf der Schale (also oben im Mündungsrand)
- Die Columellaris (Spindelwand): Der Bereich nahe der Columella (Spindel)
- Die Palatalis (Gaumenwand): Der nach außen weisende Abschnitt des Mündungsrandes.
Spindelwand und Mündungswand können unten aufeinander zulaufen und einen Kanal bilden, der bei einigen Arten recht weit ausgezogen ist. Dieser Kanal wird auch als Siphonalkanal oder Ausguss bezeichnet. Der rückwärtige (äußere) Teil des Ausgusses ist der Stiel (auch Schild genannt).
Auf dem Spindelrand können sich kleine Wülste befinden, die als Spindelfalten oder Spindelzähne bezeichnet werden. Spindelfalten können sich auch im Inneren des Gehäuses befinden, sie sind dann bei intakter Schale von außen nicht sichtbar.
Schalendeckel
Mit Hilfe eines Schalendeckels können Schnecken die Mündung ihrer Schale verschließen. Bei den Vorderkiemerschnecken handelt es sich um das so genannte Operculum, das am Fußende der Schnecke angewachsen ist. Andere Deckelformen sind das Epiphragma (hauptsächlich von der Weinbergschnecke bekannt), das temporär zum Zwecke des Schutzes gegen Frost und Austrocknung aus einem körpereigenen Sekret gebildet wird, sowie das Clausilium der Schließmundschnecken (Clausiliidae), das mit der Schale fest verbunden ist, nicht aber mit dem Weichkörper. Lediglich diese letztgenannte Form des Schalendeckels kann als Teil der Schneckenschale angesehen werden.
Paläolithische Schmuckstücke


Durchbohrte paläolithische Schneckengehäuse, die in der Türkei, in der Üçağızlı-Höhle, geborgen wurden und als Überreste von Körperschmuck („Perlenketten“) gedeutet werden, gelten als die ältesten Schmuckstücke des anatomisch modernen Menschen (Homo sapiens), die außerhalb Afrikas geborgen wurden; ihr Alter beträgt mindestens 40.000 Jahre.[3] Sie ähneln gleich alten Funden aus der Höhle Ksar Akil im Libanon. Noch deutlich älter sind die gleichfalls als Schmuck interpretierten Gehäuse von Meeresschnecken aus der Sibudu-Höhle und der Blombos-Höhle in Südafrika sowie aus der Grotte des Pigeons in Marokko.
Siehe auch
- Verarbeitung von Molluskenschalen in der Indus-Kultur
- Kaurigeld (aus Gehäusen von Kaurischnecken)
- Muschelgeld (verschiedene Molluskengelder aus Schneckengehäusen oder Muschelschalen)
Literatur
- Paul Brohmer: Fauna von Deutschland. 15. Auflage, Heidelberg 1982, ISBN 3-494-00043-3
- Andreas E. Richter: Handbuch des Fossiliensammlers. Stuttgart 1991. ISBN 3440050041
- Arno Hermann Müller: Lehrbuch der Paläozoologie, Band II, Teil 2, Jena 1981.
Weblinks
Einzelnachweise
- Hochspringen ↑ Pilsbry H. A. & Cooke C. M. 1918–1920. Manual of Conchology. Second series: Pulmonata. Volume 25. Pupillidae (Gastrocoptinae, Vertigininae). Philadelphia. page vii.
- Hochspringen ↑ Edmund Gittenberger: Beiträge zur Kenntnis der Pupillacea : III. Chondrininae. Zoologische Verhandelingen, 127(1): 3-267, 1973 ISSN 0024-1652 PDF.
- Hochspringen ↑ Richard G. Klein: The Human Career: Human Biological and Cultural Origins. 3. Auflage 2009, S. 646, ISBN 978-0-226-43965-5.