Timelife Émergence De Homme Manquant Lien Australopithecus Africanus Habilis















Série Time-Life L'émergence de l'homme – Le chaînon manquant.
NOTE: Nous avons 75 000 livres dans notre bibliothèque, soit près de 10 000 titres différents. Il y a de fortes chances que nous ayons d'autres exemplaires de ce même titre dans des conditions variables, certaines moins chères, d'autres en meilleur état. Nous pouvons également avoir différentes éditions (certaines en livre de poche, d'autres à couverture rigide, souvent des éditions internationales). Si vous ne voyez pas ce que vous voulez, veuillez nous contacter et demander. Nous sommes heureux de vous envoyer un résumé des différentes conditions et prix que nous pouvons avoir pour le même titre.
DESCRIPTION: Couverture rigide illustrée : 160 pages. Éditeur : Time-Life Books Inc. (1972).
ÉTAT : COMME NEUF à TRÈS BON. Légère usure des étagères, sinon en très bon état, comme neuf. Apparemment non lu, simplement feuilleté plusieurs fois, au pire peut-être lu une seule fois. L’intérieur des pages est vierge ; propre, non marqué, non mutilé, étroitement relié. Je voudrais seulement noter que les livres Time-Life de l'époque présentent parfois un léger « affaissement des étagères », c'est-à-dire que la reliure/les pages glissent vers le bas (peut-être 1/8 de pouce – 3 mm) à l'intérieur des couvertures au fil des ans, de sorte qu'elles finissent par reposer dessus. l'étagère. En outre, la surface supérieure des bords des pages fermées (parfois appelée « bloc de page ») présentera de légères taches de vieillesse de couleur beige – non pas sur les pages ouvertes individuelles, mais uniquement sur la surface supérieure des bords groupés des pages fermées. Si l'un ou l'autre est prononcé, nous ne proposons pas le livre – nous décrivons donc au pire un état très légèrement altéré par l'âge. Les couvertures illustrées ne témoignent que d’usures très douces sur les bords et les coins des étagères. Il n’y a aucun défaut majeur dans le livre, ni à l’intérieur ni à l’extérieur. Satisfaction garantie sans condition. En inventaire, prêt à livrer. Aucune déception, aucune excuse. EMBALLAGE LOURDEMENT REMBOURRÉ ET SANS DOMMAGE ! Des descriptions minutieuses et précises ! Vente en ligne de livres d'histoire ancienne rares et épuisés depuis 1997. Nous acceptons les retours pour quelque raison que ce soit dans les 30 jours ! #1161.
VEUILLEZ VOIR LES IMAGES CI-DESSOUS POUR LA DESCRIPTION DE LA VESTE ET POUR LES PAGES DE PHOTOS DE L’INTÉRIEUR DU LIVRE.
La série « Emergence of Man » est sortie au milieu des années 1970. Chaque volume s'engage à décrire les événements majeurs survenus au cours d'une période spécifique du développement de la ou des civilisations humaines. Les volumes sont richement illustrés et conçus comme une introduction à la période couverte. Les interprétations ou reconstitutions des artistes de ce à quoi auraient ressemblé diverses civilisations anciennes – leur architecture, leurs maisons, leurs monuments, leurs villes, leur vie quotidienne, leurs bijoux, leur nourriture, leur vie de famille, leurs habitations, leurs occupations, etc. À titre d’exemple, les ruines de Babylone et d’Ur, d’Athènes et de Rome témoignent de l’incroyable richesse de ces villes légendaires. Les reconstitutions de l'artiste dans cette série sont tout simplement ahurissantes. C’est aussi proche que possible d’y avoir été.

L’ensemble de la série est véritablement une magnifique introduction à l’histoire de l’époque. Si vous pouviez avoir un seul livre (ou une série de livres) pour présenter l’histoire de l’humanité, ce serait celui-là. Les aperçus sont concis et bien rédigés. Avec les illustrations et les images, ils donnent une merveilleuse « image » mentale et émotionnelle de ce à quoi a dû ressembler la vie dans diverses civilisations et à différentes époques. Réalisés dans un style si merveilleusement caractéristique des publications de Time-Life, ce sont des livres surdimensionnés de type « table basse » remplis d'images impressionnantes. Les images des plus grands arts et architectures du monde valent à elles seules le prix de ces livres. Mais n'ayez pas l'impression que ces volumes sont du « fluff ». Même si un volume particulier ne constitue peut-être pas tout à fait la place d'un diplôme universitaire, le matériel est bien écrit, informatif et extrêmement gratifiant intellectuellement, aussi sommaire soit-il.
Certains des sujets inclus sont énumérés ci-dessous afin de vous donner une idée de la richesse du contenu :

CONTENU:
Introduction.
CHAPITRE PREMIER : L'Ancêtre.
-La famille humaine : sera à l'homme dans 14 millions d'années.
ESSAI IMAGE : La vie quotidienne des australopithèques dans l'Éden d'un chasseur.
-Une existence liée à l'eau et au gibier.
-Clubs pour les armes, branches pour une danse de la pluie.
-La journée détendue ou un prédateur primitif.
-À la tombée de la nuit, un lieu de repos sûr dans les branches.
CHAPITRE DEUX : Les preuves des pierres et des os.
-Différences de dents : homme, singe, singe.
-Ancêtres communs des primates.
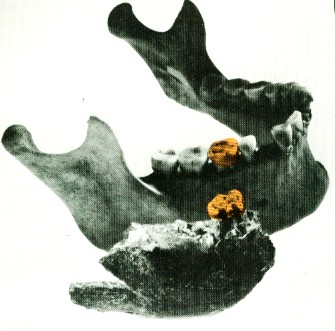
-Différences dans les mâchoires : homme, singe, singe.
-Structure squelettique : Africanus/Homo Habilis, Boisei, Homo Robustus.
-Les maigres restes d'australopithèques.
-Homo Robustus : La mâchoire Lothagam.
CHAPITRE TROIS : En bas des arbres.
-Le Lémur catta : Rappel évolutif.
-Musaraignes arboricoles : ancêtre primate ?
-Tarsier : Primate primitif.
-Potto prosimien africain : ancêtre du singe.
-Singe de velours arboricole.
-Gibbon : le singe le moins étroitement lié à l'homme.
ESSAI IMAGE : Où sont les fossiles – les sites d'hominidés célèbres d'Afrique de l'Est.
-Richard Leakey.
-Les badlands près du lac Rudolf.
-Lothagam : Source des vestiges les plus anciens.
-Omo : Une Terre Verte comme dans les Temps Anciens.
-Olduvai Gorge : site qui a révélé le chaînon manquant.

CHAPITRE QUATRE : Une niche sur la savane.
-Utilisation d'outils Primates.
-Rétroaction positive : un mécanisme évolutif.
CHAPITRE CINQ : Vie sociale des hommes-singes.
CHAPITRE SIX : Armes et outils.
-Chasse.
-Proie préhistorique.
-L'abattage du troupeau.
-Techniques de chasse en groupe.
-Partage de nourriture.
-Outils de pierre.

CHAPITRE SEPT : Nouveaux indices sur la descendance de l'homme.
-Etudes génétiques.
-Méthode ADN, Séquençage des Protéines, Immunologie.
-L'arbre généalogique : preuves dans une protéine.
-L'arbre généalogique : les fossiles réévalués.
-L'arbre généalogique : un spectre de vues.
ESSAI IMAGE : Les experts ont leur mot à dire.
Crédits, Remerciements.
Bibliographie.
Indice.

CONTEXTE SUPPLÉMENTAIRE:
LES HOMININS DE L'ÂGE DE PIERRE (PREMIÈRE PARTIE):
Homo habilis - Homo rudolfensis - Homo ergaster - Homo erectus - Homo antecessor Homo heidelbergensis - Homo neanderthalensis – Homo denisovan – Homo floresiensis - Homo sapiens.
Homo habilis: Homo habilis (« homme à tout faire ») est une espèce humaine archaïque du Pléistocène inférieur d'Afrique de l'Est et du Sud. Les vestiges sont datés d'il y a environ 2,3 à 1,65 millions d'années. La description initiale de l'espèce en 1964 a été très contestée. De nombreux chercheurs de l’époque insistaient pour que Homo habilis soit synonyme d’Australopithecus africanus. Australopithecus africanus était le seul autre hominidé ancien connu à l'époque. Cependant, Homo habilis a reçu de plus en plus de reconnaissance au fil du temps et des découvertes plus pertinentes ont été faites. Dans les années 1980, Homo habilis était considéré comme un ancêtre de l’humanité. Les chercheurs ont suggéré que l’Homo habilis a évolué directement vers l’Homo erectus, ce qui a directement conduit à l’homme moderne.
Ce point de vue est désormais controversé. Plusieurs spécimens dont l'identification n'est pas sûre ont été attribués à Homo habilis, ce qui a conduit à des arguments en faveur de la division des spécimens en "Homo rudolfensis" et "Homo gautengensis". Seule la classification d'Homo rudolfensis a reçu un large soutien de la part des chercheurs. La taille du cerveau de l'Homo habilis variait généralement entre 500 et 900. Les proportions corporelles d'Homo habilis ne sont connues qu'à partir de deux squelettes extrêmement fragmentaires. Les calculs reposent donc en grande partie sur l’hypothèse d’une anatomie similaire à celle des australopithèques antérieurs. Pour cette raison, de nombreux chercheurs ont proposé que Homo habilis soit déplacé vers le genre Australopithecus sous le nom d'Australopithecus habilis.
L'interprétation d'Homo habilis comme un être humain de petite taille doté de capacités de déplacement inefficaces sur de longues distances a été remise en question. Un spécimen femelle présumé est traditionnellement interprété comme mesurant entre 3 pieds 3 pouces et 3 pieds 11 pouces de hauteur et entre 44 et 82 livres de poids. Cependant, si des proportions plus humaines étaient utilisées comme base pour ces calculs, les dimensions ressembleraient davantage à 4 pieds 10 pouces de hauteur et à environ 77 livres de poids. Quelles que soient ses dimensions, Homo habilis pourrait avoir été au moins partiellement arboricole, comme ce qui est postulé pour les australopithèques.
Les premiers hominidés sont généralement reconstruits comme ayant des cheveux épais et un dimorphisme sexuel marqué, les mâles étant beaucoup plus gros que les femelles. Ceci malgré le fait que la taille relative des hommes et des femmes est spéculative et n’est pas définitivement connue. Homo habilis fabriquait des outils en pierre de l'industrie Oldowan, utilisant principalement les pierres comme aides au dépeçage des carcasses d'animaux. Par rapport aux australopithèques, on pense généralement que les premières espèces d’Homo ont consommé de grandes quantités de viande. Dans le cas d'Homo habilis, on pense qu'il s'agissait principalement de viande récupérée.
Généralement, les premiers hominidés sont interprétés comme ayant vécu dans des sociétés polygames. Mais cela aussi est hautement spéculatif. En supposant que la société Homo habilis était similaire à celle des chimpanzés et des babouins des savanes modernes, les groupes pouvaient compter entre 70 et 85 membres. Plusieurs mâles se seraient défendus contre les prédateurs de la savane ouverte, tels que les félins, les hyènes et les crocodiles. Homo habilis coexistait avec Homo rudolfensis, Homo ergaster et/ou Homo erectus et Paranthropus boisei.
Les premiers restes reconnus d'Homo habilis étaient des os partiels de crâne, de main et de pied juvénile datant d'il y a 1,75 million d'années. Ils ont été découverts en 1960 dans les gorges d'Olduvai, en Tanzanie, par Jonathan Leakey. Cependant, des restes non reconnus sont antérieurs à ceux-ci. Les premiers restes, constitués d'une molaire, ont été découverts par l'assistant principal de Louis et Mary Leakey (les parents de Jonathan) en 1959. Cependant, on ne se rendait pas compte à l'époque que la molaire appartenait à une nouvelle espèce, Homo habilis. Au moment de la découverte de 1959, Louis et Mary Leakey avaient passé 29 ans à fouiller dans les gorges d'Olduvai à la recherche des premiers restes d'hominidés, mais avaient plutôt récupéré principalement d'autres restes d'animaux ainsi que des outils en pierre témoignant de l'industrie des outils en pierre d'Oldowan.
L'industrie des outils en pierre d'Oldowan avait été attribuée à Paranthropus boisei, connu à l'époque sous le nom de « Zinjanthropus ». En 1959, Paranthropus boisei fut le premier et le seul hominidé découvert dans la région. Mais cette situation a bien sûr changé avec la découverte de fossiles attribuables à Homo habilis. En 1964, les paléoanthropologues et primatologues ont officiellement classé les restes dans le genre Homo. Sur recommandation des anthropologues, le nom spécifique Homo habilis. Homo habilis signifiait « capable, maniable, mentalement habile, vigoureux » en latin. À cette époque, l’industrie des outils en pierre d’Oldowan était considérée comme une preuve de capacités cognitives avancées. L'association de l'Homo habilis avec l'Oldowan a également été utilisée comme justification pour le classer dans Homo.
Après avoir été classé, les chercheurs ont débattu vivement pour savoir si Homo habilis devait être reclassé en Australopithecus africanus. Cela était en partie dû au fait que les restes étaient si vieux et qu'à l'époque, Homo était présumé avoir évolué en Asie. De plus, Australopithecus africanus était le seul autre hominidé ancien connu à l'époque, et les australopithèques n'avaient pas de descendants. De plus, la taille du cerveau d'Homo habilis était inférieure à ce que les paléoanthropologues proposaient en 1955 comme minimum pour les espèces Homo. Cependant, la classification Homo habilis a commencé à être plus largement acceptée à mesure que davantage d'éléments et d'espèces fossiles ont été découverts.
En 1983, des paléoanthropologues ont proposé qu'Australopithecus africanus soit un ancêtre direct de Paranthropus et d'Homo (les deux étaient des populations sœurs). En outre, ils ont proposé que l'Australopithecus africanus ait évolué vers Homo habilis, qui à son tour a évolué vers Homo erectus. La maison erectus a à son tour évolué vers l’homme moderne. Toutes ces évolutions ont été attribuées à un processus de cladogenèse, ou de détournement/scission. Une autre proposition était qu'il y avait un saut évolutif majeur entre Australopithecus africanus et Homo habilis. Par la suite, il a été proposé que l’évolution humaine ait progressé progressivement parce que la taille du cerveau d’Homo habilis avait déjà presque doublé par rapport à ses prédécesseurs australopithèques.
Cependant, à cette époque, le plus ancien spécimen d'Homo habilis était très similaire aux spécimens plus jeunes, ce qui impliquait qu'il n'y avait pas de progression évolutive entre deux spécimens largement espacés. De nombreux paléoanthropologues avaient accepté le modèle proposé. Les restes d'hominidés de la fin du Pliocène au début du Pléistocène en dehors de l'aire de répartition du Paranthropus et de l'Homo erectus avaient été classés dans l'Homo habilis. Pour les éléments autres que le crâne, cela a été fait sur la base de la taille, car il manquait des caractéristiques diagnostiques claires. Grâce à cette pratique, la gamme de variations de l'espèce est devenue assez large. Les termes Homo habilis sensu stricto (« au sens strict ») et Homo habilis sensu lato (« au sens large ») étaient utilisés pour inclure et exclure, respectivement, les formes les plus divergentes.
Pour répondre à cette grande variation des caractéristiques d'Homo habilis, les paléoanthropologues suggèrent en 1985 l'existence d'une espèce différentielle. Le crâne relativement massif du lac Turkana, au Kenya, découvert en 1972, avait été attribué à Homo habilis. Les paléoanthropologues ont suggéré que ces fossiles représentaient en réalité une espèce différente, maintenant connue sous le nom d'Homo rudolfensis (le lac Turkana était autrefois connu sous le nom de lac Rudolf). Au contraire, d'autres paléoanthropologues ont soutenu que plutôt qu'une nouvelle espèce, le plus grand spécimen du lac Turkana était un spécimen mâle, alors que les autres spécimens d'Homo habilis étaient des femelles. Le dimorphisme sexuel expliquerait la variation de taille au sein d'une même espèce.
Ces paléoanthropologues ont suggéré que tous les premiers Homo d'Afrique du Sud peuvent être attribués aux classifications Homo habilis ou Homo ergaster et/ou Homo erectus. Cependant, en 2010, les archéologues ont proposé de diviser les premiers Homo sud-africains en une nouvelle espèce, Homo gautengensis. En 1986, un squelette fragmentaire avait été découvert par des anthropologues américains en association avec des fragments de crâne d'Homo habilis. Pour la première fois, des aspects de l’anatomie squelettique d’Homo habilis ont été définitivement établis. Les restes squelettiques ont révélé des caractéristiques plus proches de celles d'Australopithèque que d'Homo.
En raison de cette révélation ainsi que des similitudes dans les caractéristiques dentaires, les paléoanthropologues ont suggéré en 1999 de reclasser Homo habilis en Australopithèque. Cependant, une réévaluation ultérieure d'une physiologie plus humaine, si elle était correcte, jetterait le doute sur ce point. Puis vint la découverte des crânes géorgiens de Dmanisi vieux de 1,8 million d’années au début des années 2000. Ces crânes présentent plusieurs similitudes avec les premiers Homo. Cela a conduit les paléoanthropologues à suggérer que tous les groupes contemporains des premiers Homo en Afrique sont de la même espèce et devraient être reclassés comme Homo erectus. Cela comprendrait Homo habilis et Homo rudolfensis.
Il n’existe toujours pas de large consensus quant à savoir si Homo habilis est ou non l’ancêtre de Homo ergaster et/ou de Homo erectus. La question de savoir si l'Homo habilis est une émanation de la lignée humaine fait l'objet d'un vif débat. Il est également contesté si tous les spécimens attribués à Homo habilis sont correctement attribués. Certains soutiennent qu'Homo habilis n'est rien de plus qu'un assemblage de différentes espèces d'Australopithèques et d'Homo. Néanmoins, Homo habilis et Homo rudolfensis sont généralement des membres reconnus du genre à la base de l'arbre généalogique Homo. Les arguments en faveur de la synonymisation ou du retrait d'Homo habilis du genre ne sont pas largement adoptés.
Bien qu'il soit désormais largement admis qu'Homo a évolué à partir de l'Australopithèque, le moment et le lieu de cette scission ont fait l'objet de nombreux débats. De nombreuses espèces d'australopithèques différentes ont été proposées comme ancêtre d'Homo habilis. Le plus ancien spécimen d’Homo jamais découvert date d’il y a 2,8 millions d’années. L’espèce à laquelle cette plus ancienne découverte d’Homo est attribuable n’est pas claire. Il a été trouvé dans la région Afar d'Éthiopie. Il pourrait s'agir de l'ancêtre d'Homo habilis, mais cela reste incertain. Cependant, la découverte pourrait indiquer qu'Homo habilis pourrait avoir évolué à partir d'Australopithecus afarensis à cette époque.
Le spécimen le plus ancien positivement classé comme Homo habilis date d'il y a 2,3 millions d'années. Cependant, le spécimen est anatomiquement plus dérivé. Il possède moins de traits ancestraux ou basaux que les spécimens plus jeunes. Cela suggère que les formes dérivées et basales ou leurs variations vivaient simultanément. Cela confirmerait que la lignée d’Homo habilis a commencé il y a 2,3 millions d’années. D'après des outils en pierre vieux de 2,1 millions d'années provenant de Shangchen, en Chine, Homo habilis ou une espèce ancestrale pourrait s'être dispersée à travers l'Asie. Le plus jeune spécimen d’Homo habilis date d’il y a environ 1,65 million d’années.
On pense généralement que la taille du cerveau augmente le long de la lignée humaine, particulièrement rapidement lors de la transition entre les espèces. La taille du cerveau d'Homo habilis était plus petite que celle des Homo ergaster et/ou Homo erectus qui lui succédèrent. La taille du cerveau est passée d'environ 600 à 650 cc chez Homo habilis à environ 900 à 1 000 cc chez Homo ergaster et Homo erectus. Cependant, une étude de 2015 a montré que la taille du cerveau d'Homo habilis, Homo rudolfensis et Homo ergaster se situait généralement entre 500 et 900 cc. La taille du cerveau des australopithèques variait généralement entre 400 et 500 cc. L'anatomie cérébrale de tous les Homo présente un cerveau élargi par rapport aux australopithèques.
Un motif de stries inclinées vers la droite sur les dents d'un spécimen d'Homo habilis peut avoir été accidentellement auto-infligé. L'individu aurait pu tirer un morceau de viande avec ses dents et la main gauche, tout en essayant de le couper avec un outil en pierre en utilisant la main droite. Si cela est exact, cela pourrait indiquer une droiterie, associée à une réorganisation majeure du cerveau et à la latéralisation de la fonction cérébrale entre les hémisphères gauche et droit. Ce scénario a également été émis pour certains spécimens de Néandertal.
La latéralisation pourrait être impliquée dans l’utilisation des outils. Chez l’homme moderne, la latéralisation est faiblement associée au langage. Les rangées de dents d'Homo habilis étaient en forme de V, par opposition à la forme de U chez Homo plus tard. La bouche faisait saillie vers l'extérieur, bien que le visage soit plat du nez vers le haut. Basée sur des squelettes fragmentaires d'individus que l'on croit être des hommes et des femmes, l'anatomie corporelle d'Homo habilis a généralement été considérée comme ressemblant davantage à un singe que même celle de l'Australopithecus afarensis antérieur. Cela serait cohérent avec un mode de vie au moins partiellement arboricole dans les arbres, comme on le suppose chez les australopithèques.
Sur la base des restes squelettiques fragmentaires décrits ci-dessus et en supposant des dimensions corporelles comparables à celles des australopithèques, Homo habilis a généralement été interprété comme ayant été de petite taille comme les australopithèques. L'Homo habilis typique est généralement estimé à environ 3 pieds, 3 pouces à 3 pieds, 11 pouces de hauteur et 44 à 82 livres de poids. Cependant, en supposant des jambes humaines plus longues et modernes, la hauteur pourrait être plutôt de 4 pieds 10 pouces et le poids moyen d'environ 77 livres. En comparaison, les hommes et les femmes modernes en 1900 mesuraient en moyenne respectivement 5 pieds, 4 pouces et 5 pieds.
On suppose généralement que les hominidés pré-Homo ergaster, y compris Homo habilis, présentaient un dimorphisme sexuel notable. Si cette hypothèse est vraie, les mâles seraient nettement plus gros que les femelles. Cependant, il ne s’agit que d’une simple hypothèse, car la masse corporelle relative des femelles est inconnue chez cette espèce. On pense que Homo habilis, comme les autres premiers hominidés, avait une épaisse couverture de poils sur le corps, comme les singes non humains modernes. Cela est dû en partie au fait qu’ils semblent avoir habité des régions plus froides. On pense également qu’ils ont eu un mode de vie moins actif que les espèces post-ergaster (présumées glabres). Dans ces circonstances, ils avaient probablement besoin de poils épais pour rester au chaud.
Sur la base des taux de développement dentaire, on suppose qu'Homo habilis a eu un taux de croissance accéléré par rapport à l'homme moderne. Le taux de croissance imputé aurait été plus proche de celui des singes non humains modernes. Les bras de l'Homo habilis et des australopithèques sont généralement considérés comme étant proportionnellement longs et donc adaptés à l'escalade et au balancement. Cependant, le rapport humérus/fémur de nombreux spécimens d'Homo habilis et d'Australopithecus afarensis se situe dans la plage de variation de l'homme moderne, il peut donc être dangereux de prendre des proportions semblables à celles d'un singe. Néanmoins, le rapport humérus/fémur de certains spécimens se situe en dehors de la plage de variation des humains modernes, plus proche de la proportion observée chez les chimpanzés.
Les os de la main d'Homo habilis suggèrent une préhension précise, importante pour la dextérité, ainsi que des adaptations pour l'escalade. En ce qui concerne le fémur, des comparaisons traditionnelles avec les australopithèques ont été utilisées pour reconstruire des pattes robustes chez Homo habilis. Cependant, le fémur plus gracile d'Homo ergaster et/ou d'Homo erectus ou de Paranthropus boisei peut constituer une comparaison plus appropriée. Si tel était le cas, Homo habilis aurait eu des jambes plus longues, semblables à celles des humains. Homo habilis aurait été un voyageur efficace sur de longues distances, comme cela aurait été le cas pour Homo ergaster. Cependant, l'estimation de la longueur non préservée d'un fossile est très problématique.
L'épaisseur des os des membres de certains spécimens d'Homo habilis ressemble plus à celle des chimpanzés qu'à celle de l'Homo ergaster et/ou de l'Homo erectus et des humains modernes. Cela peut indiquer des capacités portantes différentes, plus adaptées à l'arboréalité chez Homo habilis. Le péroné solide ressemble davantage à celui des singes non humains et est cohérent avec l'arboréalité et l'escalade verticale. Les pieds sont mieux adaptés aux mouvements terrestres que ceux de l'Australopithecus afarensis, bien qu'ils conservent de nombreuses caractéristiques simiesques compatibles avec l'escalade. Cependant, le pied présente des structures articulaires saillantes et compactées au milieu du pied, qui limitent la rotation entre le pied et la cheville ainsi qu'à l'avant du pied.
La stabilité du pied améliore l'efficacité du transfert de force entre la jambe et le pied et vice versa. C'est un composant essentiel du mécanisme à ressort élastique de la voûte plantaire qui génère de l'énergie pendant la course (mais pas la marche). Cela pourrait éventuellement indiquer qu'Homo habilis était capable d'un certain degré de course d'endurance. On pense généralement que cette caractéristique a évolué plus tard chez Homo ergaster et/ou Homo erectus. Les preuves suggèrent qu'il pourrait s'être développé plus tôt dans l'évolution d'Homo.
Traditionnellement, Homo ergaster et/ou Homo erectus est considéré comme le premier humain à avoir vécu dans une société monogame. La plupart des chercheurs pensent que tous les hominidés précédents étaient polygames. Cependant, il est extrêmement difficile de spéculer avec certitude sur la dynamique de groupe des premiers hominidés. Le problème réside dans le degré de dimorphisme sexuel et la disparité de taille entre les mâles et les femelles. Sur cette seule base, les chercheurs établissent une corrélation entre la polygynie avec une forte disparité et la monogamie avec une faible disparité, en se basant sur les tendances générales observées chez les primates modernes. Il existe quelques exceptions à cette corrélation. Mais d’une manière générale, il s’agit là d’une description précise de la méthodologie utilisée pour spéculer sur la prévalence générale de la monogamie.
Cependant, la détermination des taux de dimorphisme sexuel est problématique et incertaine en raison du fait que l’anatomie précoce des hominidés n’est pas bien établie. Le peu que l’on sait repose en grande partie sur quelques spécimens seulement. Dans certains cas, le sexe attribuable aux restes repose en grande partie sur la taille perçue et la robustesse apparente. Cette méthodologie est employée en l'absence d'éléments plus fiables dans l'identification du sexe (à savoir le bassin).
Les comportements d'accouplement imputés sont également basés sur l'anatomie dentaire. Mais les premiers hominidés possèdent une anatomie en mosaïque de différents traits que l’on ne retrouve pas ensemble chez les primates modernes. Les dents des joues élargies suggéreraient un dimorphisme marqué lié à la taille. Ainsi, le comportement inclurait d’intenses conflits entre hommes concernant les partenaires et une société polygame. Cependant, par la même mesure, les petites canines devraient indiquer le contraire. D’autres pressions sélectives, notamment l’alimentation, peuvent également avoir un impact considérable sur l’anatomie dentaire. L’anatomie dentaire est d’une fiabilité douteuse dans la détermination des comportements sociaux.
La répartition spatiale des outils et des os d'animaux transformés sur les sites d'Homo habilis dans les gorges d'Olduvai indique que les habitants utilisaient cette zone comme terrain de boucherie et de restauration communal du groupe. Cela contrasterait avec le système familial nucléaire des chasseurs-cueilleurs modernes où le groupe est subdivisé en unités familiales unifamiliales. Chaque famille possède son propre espace de « boucherie » et de restauration (cuisines et salles à manger). Le comportement des premiers Homo est parfois calqué sur celui des chimpanzés des savanes et des babouins. Cette modélisation inclurait Homo habilis.
Ces communautés sont composées de plusieurs hommes (par opposition à une société de harem). Ces mâles défendent le groupe sur l'habitat dangereux et exposé. Ils se livrent parfois à une démonstration de groupe de bâtons de jet et de pierres contre les ennemis et les prédateurs. Parmi les fossiles d'Homo habilis découverts, le pied gauche de l'un d'entre eux semble avoir été mordu par un crocodile, peut-être Crocodylus anthropophagus. La patte d'un autre spécimen d'Homo habilis (ou peut-être de Paranthropus) montre des preuves de prédation par le léopard. Homo habilis et les hominidés contemporains étaient probablement la proie d'autres grands carnivores de l'époque. Ceux-ci auraient inclus dans les gorges d'Olduvai l'hyène chasseuse Chasmaporthetes nitidula, ainsi que les chats à dents de sabre Dinofelis et Megantereon.
En 1993, une étude menée par des paléoanthropologues et des psychologues évolutionnistes a estimé que la taille du groupe Homo habilis variait entre 70 et 85 membres. Cela les placerait à l’extrémité supérieure de la taille des groupes de chimpanzés et de babouins. Leurs estimations étaient basées sur les tendances observées dans la taille du néocortex et la taille des groupes chez les primates non humains modernes. Homo habilis coexistait avec Homo rudolfensis, Homo ergaster et/ou Homo erectus et Paranthropus boisei. On ne sait pas exactement comment toutes ces espèces ont interagi. On pense généralement que le régime alimentaire de l’Homo habilis et des autres premiers Homo comportait une plus grande proportion de viande que celle des Australopithèques, et que cela conduisait à la croissance du cerveau. Les principales hypothèses à ce sujet sont les suivantes : la viande est riche en énergie et en nutriments. Cela a exercé une pression évolutive sur le développement de compétences cognitives améliorées pour faciliter la récupération stratégique et monopoliser les carcasses fraîches.
La viande a permis à l’intestin du singe, gros et coûteux en calories, de diminuer en taille. Cela a permis à l’énergie autrement dépensée dans l’intestin d’être détournée vers la croissance du cerveau. Alternativement, il est également suggéré que le premier Homo s'adaptait à un climat asséché avec des options alimentaires plus rares. L'adaptation devait s'appuyer principalement sur les organes de stockage souterrains (comme les tubercules) et le partage de nourriture. Cela a facilité les liens sociaux entre les membres masculins et féminins du groupe. Cependant, les premiers Homo de petite taille sont généralement considérés comme incapables de courir et de chasser en endurance. Les attributs étaient des capacités imputées à Homo ergaster et plus tard à Homo, et non à Homo habilis.
On pense que la chasse et la cueillette organisées ont émergé chez Homo ergaster. L'avant-bras long et semblable à un australopithèque d'Homo habilis pourrait indiquer que les premiers Homo étaient encore arboricoles dans une certaine mesure. Le problème est alors que les modèles de collecte de nourriture proposés, qui seraient nécessaires pour expliquer une croissance cérébrale importante, nécessiteraient une distance de déplacement quotidienne accrue. Ceci est contredit par la physiologie généralement imputée par les paléoanthropologues à Homo habilis à partir de restes. Cependant, certains paléontologues ont soutenu qu'Homo habilis possédait peut-être des jambes humaines modernes. Cela signifierait que Homo habilis aurait pu être pleinement capable de voyager efficacement sur de longues distances, tout en restant au moins partiellement arboricole.
La grande taille des incisives chez Homo habilis par rapport aux prédécesseurs de l'Australopithecus implique que cette espèce s'appuyait davantage sur les incisives. Les corps des mandibules d'Homo habilis et d'autres premiers Homo sont plus épais que ceux des humains modernes et de tous les singes vivants. Les mandibules sont plus comparables à celles des australopithèques. Le corps mandibulaire résiste à la torsion due à la force de morsure ou à la mastication. Cela signifie que les mâchoires d’Homo habilis pourraient produire un stress inhabituellement puissant en mangeant. Le plus grand relief des cuspides molaires chez Homo habilis par rapport à l'Australopithèque suggère que les premiers utilisaient des outils pour briser les aliments durs tels que les parties de plantes souples ou la viande). Si cela n'avait pas été le cas, les cuspides des molaires présenteraient une usure plus importante que celle constatée dans les archives fossiles. Néanmoins, les adaptations de la mâchoire pour le traitement d’aliments mécaniquement difficiles indiquent que les progrès technologiques n’ont pas eu d’effet prononcé sur l’alimentation.
On pense qu’Homo habilis tirait sa viande de la chasse plutôt que de la chasse. Le rôle attribué par les paléontologues fait qu'Homo habilis agit comme un charognard conflictuel et vole les proies de petits prédateurs tels que les chacals ou les guépards. Les fruits constituaient probablement également un élément important du régime alimentaire. Ceci est indiqué dans les archives fossiles par une érosion dentaire compatible avec une exposition répétée à l'acidité. D’après l’analyse de la texture des micro-usures dentaires, Homo habilis ne consommait probablement pas régulièrement d’aliments durs. Ceci est cohérent avec les archives fossiles laissées par d’autres premières espèces d’Homo. En moyenne, la complexité de la texture des micro-usures d'Homo habilis se situe quelque part entre celle des mangeurs de nourriture coriaces et celle des mangeurs de feuilles (folivores). Cela témoigne d’une alimentation de plus en plus généralisée et omnivore.
Homo habilis est associé à l’industrie des outils en pierre Oldowan du début de l’âge de pierre. Les individus utilisaient probablement ces outils principalement pour massacrer, écorcher des animaux et broyer des os. Cependant, il se peut que les mêmes outils aient parfois été utilisés pour gratter du bois et couper des plantes tendres. Les tailleurs semblent avoir des noyaux lithiques soigneusement sélectionnés. Homo habilis savait évidemment que certaines roches se briseraient d'une manière spécifique lorsqu'elles seraient frappées assez fort et au bon endroit. Homo habilis produisait plusieurs types différents d'outils en pierre. Ceux-ci comprenaient des choppers, des polyèdres et des discoïdes. Il est néanmoins peu probable que des formes spécifiques aient été conçues à l'avance. Ils proviennent probablement simplement d’un manque de standardisation dans la production de tels outils. Les types différents de matières premières dont disposaient les tailleurs auraient également contribué à la variation de la forme des outils.
Par exemple, les sphéroïdes sont courants à Olduvai, qui présente une abondance de gros morceaux mous de quartz et de quartzite. D'un autre côté, Koobi Fora manque de sphéroïdes et fournit principalement des roches de lave basaltiques dures. Contrairement à la culture acheuléenne ultérieure inventée par Homo ergaster et/ou Homo erectus, la technologie Oldowan ne nécessite pas de planification ni de prévoyance pour sa fabrication. Par conséquent, la possession de la technologie n'indique pas une cognition élevée chez les tailleurs d'Oldowan. Cela nécessite un certain degré de coordination et des connaissances en mécanique. Les outils Oldowan présentent rarement des retouches et étaient probablement pour la plupart jetés immédiatement après utilisation.
La technologie des outils en pierre Oldowan a été signalée pour la première fois en 1934. Cependant, ce n'est que dans les années 1960 qu'elle est devenue largement acceptée comme la culture la plus ancienne et que les outils en pierre Oldowan ont été fabriqués par Homo habilis. La technologie/culture était datée d’il y a 1,8 million d’années. Depuis les années 1960, d'autres découvertes ont placé les origines de la culture matérielle considérablement en arrière dans le temps. Des outils en pierre Oldowan ont été découverts à Ledi-Geraru et Gona en Éthiopie il y a 2,6 millions d'années. La culture/technologie peut-être associée à l'évolution du genre.
Les australopithèques sont également connus pour avoir fabriqué des outils, comme l'industrie des outils en pierre de Lomekwi, vieille de 3,3 millions d'années. Il existe également des preuves de dépeçage datant d'environ 3,4 millions d'années. Néanmoins, la culture Oldowan, relativement pointue, constituait une innovation majeure par rapport à la technologie australopithèque. Cela aurait permis différentes stratégies d'alimentation et la capacité de traiter une plus large gamme d'aliments. Cela aurait été très avantageux dans le climat changeant de l’époque. On ne sait pas si l’industrie des outils en pierre d’Oldowa a été inventée de manière indépendante ou si elle est le résultat d’expérimentations d’hominidés avec des roches sur des centaines de milliers d’années et sur plusieurs espèces.
En 1962, un cercle de 12 pieds × 14 pieds × 1 pied fait de roches volcaniques a été découvert dans les gorges d'Olduvai. À des intervalles de 2 à 2 pieds et demi, les roches étaient empilées sur une hauteur de 6 à 9 pouces. Mary Leakey a suggéré que les tas de roches étaient utilisés pour soutenir des poteaux enfoncés dans le sol. Cela aurait pu être fait pour soutenir un brise-vent ou une cabane rudimentaire. Certaines tribus nomades des temps modernes construisent des parois rocheuses basses similaires pour y construire des abris temporaires. Les cercles rocheux sont utilisés pour contenir des branches verticales courbées utilisées comme poteaux. Des herbes ou des peaux d'animaux étaient ensuite placées au sommet de la structure des poteaux courbés comme écran de couverture. Datant d'il y a 1,75 million d'années, il est attribué à un genre Homo ancien et constitue la plus ancienne preuve d'architecture découverte à ce jour [Wikipedia].
Homo rudolfensis: Homo rudolfensis est une espèce humaine archaïque du Pléistocène inférieur d'Afrique de l'Est datée d'il y a environ 2 millions d'années. Homo rudolfensis coexistait avec plusieurs autres hominines. Par conséquent, il y a un débat sur les spécimens qui peuvent être attribués avec confiance à cette espèce au-delà de quelques crânes découverts. Aucun reste corporel n'est définitivement attribué à Homo rudolfensis. Par conséquent, sa classification générique et sa validité sont débattues sans un large consensus. Certains recommandent même que l'espèce appartienne réellement au genre Australopithecus comme A. rudolfensis ou Kenyanthropus comme K. rudolfensis, ou qu'elle est synonyme de l'Homo habilis contemporain et anatomiquement similaire.
Homo rudolfensis se distingue de Homo habilis par sa plus grande taille. Cependant, certains paléoanthropologues affirment que cette espèce est en réalité constituée de spécimens mâles d'Homo habilis. Cela supposerait que Homo habilis était sexuellement dimorphe et que les mâles étaient beaucoup plus gros que les femelles. Puisqu'aucun reste corporel n'est définitivement identifié, les estimations de la taille du corps sont largement basées sur la stature de l'Homo habilis. En utilisant ces estimations, Homo rudolfensis pourrait mesurer en moyenne environ 5 pieds 3 pouces de hauteur et peser environ 132 livres. Les femelles peuvent mesurer en moyenne environ 4 pieds 11 pouces et peser environ 112 livres.
Un fossile de crâne vraisemblablement représentatif avait un volume cérébral d'environ 750 cc. Comme les autres premières espèces d'Homo, Homo rudolfensis avait de grandes dents jugales et un émail épais. Les premières espèces d'Homo présentent une croissance cérébrale marquée par rapport aux prédécesseurs des Australopithèques. Les paléoarchéologues expliquent généralement que cela est dû à un changement de régime alimentaire avec une source alimentaire riche en calories, à savoir la viande. Bien qu’elle ne soit pas associée aux outils, l’anatomie dentaire suggère un certain traitement des fibres végétales ou carnées avant consommation. Néanmoins, les structures robustes de la bouche pouvaient toujours mâcher efficacement des aliments mécaniquement difficiles, ce qui indique que l’utilisation d’outils n’affectait pas beaucoup le régime alimentaire.
Les premiers fossiles d'Homo rudolfensis ont été découverts en 1972 au bord du lac Turkana au Kenya. À l’époque, le lac Turkana s’appelait lac Rudolf au Kenya, d’où le nom d’Homo rudolfensis. Les fossiles ont été détaillés par le paléoanthropologue kenyan Richard Leakey l'année suivante. Les spécimens consistaient en un crâne large et presque complet ; un fémur droit ; un fragment de fémur supérieur (proximal) ; et un fémur gauche complet. Les quatre fossiles ont été découverts par trois individus différents. Et on ne sait pas vraiment si les fossiles des fémurs appartenaient à la même espèce que le crâne. Leakey a classé tous les fossiles sous le genre Homo parce qu'il avait reconstruit les fragments de crâne de manière à ce qu'ils aient un volume cérébral important et une face plate. Cependant, les fossiles n’étaient pas attribués à une espèce à cette époque.
Parce que l’horizon dans lequel ils ont été découverts était daté d’il y a 2,9 à 2,6 millions d’années, Leakey pensait que ces spécimens étaient un très premier ancêtre humain. Cela remettait en question le modèle majeur de l’évolution humaine de l’époque. Si Australopithecus africanus avait donné naissance à Homo il y a environ 2,5 millions d'années, mais qu'Homo existait déjà à cette époque, cela nécessiterait de sérieuses révisions. Cependant, en 1977, une analyse plus approfondie a daté l'horizon de la région à environ 2 millions d'années. Cela a coïncidé avec la même période que Homo habilis et Homo ergaster/Homo erectus. Une analyse plus approfondie en 2012 a fourni une datation plus précise du matériau fossile, il y a 2,1 à 1,95 millions d'années.
Les fossiles ont été classés pour la première fois par les anthropologues comme Homo habilis en 1975. En 1978, Leakey a suggéré que les restes devraient être classés comme australopithèques, bien qu'il ait admis que les restes pourraient appartenir à Homo habilis. Cependant, le crâne n'a pas été correctement reconstruit. En 1986, des paléoanthropologues ont suggéré que les restes représentaient une espèce Homo différente. Ils ont suggéré que les espèces encore inconnues coexistaient avec Homo habilis et Homo ergaster/Homo erectus. Les espèces Homo coexistantes étaient en conflit avec le modèle prédominant de l’évolution humaine à l’époque, selon lequel les humains modernes évoluaient directement en ligne droite. L’opinion prédominante était que les humains modernes ont évolué directement à partir de l’Homo ergaster/Homo erectus, qui ont eux-mêmes évolué directement à partir de l’Homo habilis.
En 1986, les restes ont été classés comme une nouvelle espèce, rudolfensis. Cependant la validité de cette espèce est débattue. Certains paléoanthropologues soutiennent qu'Homo habilis était fortement dimorphique sexuellement, tout comme les singes non humains modernes. Ils pensent que les plus gros crânes classés comme « Homo rudolfensis » représentent en réalité un Homo habilis mâle. En 1999, les anthropologues biologiques ont recommandé de reclasser Homo rudolfensis et Homo habilis en Australopithèque (non Homo) sur la base de la similitude des adaptations dentaires. Cependant, ils ont admis que l'anatomie dentaire est très variable parmi les hominidés et n'est pas toujours fiable lors de la formulation d'arbres généalogiques.
En 2003, les anthropologues ont conclu que la classification antérieure des australopithèques Kenyanthropus platyops par Leakey était l'ancêtre de rudolfensis et l'ont reclassé sous le nom de K. rudolfensis. Ils ont exprimé leur conviction que Kenyanthropus était plus étroitement lié à Paranthropus qu'à Homo. En 2008, une reconstruction du crâne a conclu qu'il n'avait pas été correctement restauré à l'origine. Néanmoins, l'équipe de paléoanthropologues était d'accord avec la classification comme Homo rudolfensis. En 2012, le paléoanthropologue britannique Meave Leakey a décrit un visage partiel juvénile découvert à Koobi Fora, au Kenya, comme étant Homo rudolfensis.
Elle a noté que le crâne plus petit partageait plusieurs similitudes avec un autre crâne plus grand. Elle l'a classé comme Homo rudolfensis parce que les os prépubères des hommes et des femmes sont pratiquement impossibles à distinguer. Les différences entre les spécimens juvéniles d'Homo rudolfensis et les adultes d'Homo habilis soutiennent la distinction des espèces. Elle a également conclu qu’une mâchoire classée comme Homo rudolfensis appartient en réalité à une espèce différente, peut-être non décrite. Cependant, d'autres paléoanthropologues estiment que l'introduction d'une autre espèce est prématurée car on ne sait pas exactement quelle est l'étendue de la variation chez les premiers hominidés.
L'année 2013 a été marquée par la découverte de crânes de Dmanisi vieux de 1,8 million d'années, provenant du pays eurasien de Géorgie. Ces crânes présentent plusieurs similitudes avec les premiers Homo. Cela a conduit les chercheurs à suggérer que tous les groupes contemporains des premiers Homo en Afrique, y compris Homo habilis et Homo rudolfensis, sont de la même espèce et devraient être attribués à Homo erectus. Cela démontre que jusqu'à aujourd'hui, il n'existe toujours pas de large consensus sur la relation entre rudolfensis et habilis et Homo ergaster et les espèces descendantes. À l'exception du crâne presque intact, il existe un désaccord sur les spécimens qui appartiennent réellement à la catégorie Homo rudolfensis. Il est très difficile d’attribuer avec précision une classification aux restes qui ne préservent pas le visage et la mâchoire.
Aucun élément corporel d'Homo rudolfensis n'a jamais été définitivement associé à un crâne et donc à l'espèce. La plupart des fossiles d'Homo rudolfensis proposés proviennent de Koobi, au Kenya, et datent d'il y a 1,9 à 1,85 million d'années. Les vestiges de la formation de Shungura, en Éthiopie, et d'Uraha, au Malawi, remontent à 2,5 à 2,4 millions d'années, ce qui en ferait la première espèce d'Homo identifiée. Le dernier spécimen potentiel, Homo rudolfensis, est daté d'il y a 1,65 à 1,55 million d'années.
Néanmoins, Homo rudolfensis et Homo habilis sont généralement des membres reconnus du genre à la base de l'arbre généalogique. Les arguments en faveur de la synonymisation ou du retrait du genre ne sont pas largement soutenus. Bien qu'il soit désormais largement admis qu'Homo a évolué à partir de l'Australopithèque, le moment et le lieu de cette scission ont fait l'objet de nombreux débats. De nombreuses espèces d'australopithèques différentes ont été proposées comme ancêtre d'Homo rudolfensis et d'Homo habilis. Le plus ancien spécimen d’Homo jamais découvert date d’il y a 2,8 millions d’années. Les fossiles ont été trouvés dans la région Afar d’Éthiopie.
Ils peuvent indiquer que le genre Homo a évolué à partir d'Australopithecus afarensis à cette époque, et qu'ils pourraient être l'ancêtre d'Homo rudolfensis et d'Homo habilis. Mais l'hypothèse n'est pas certaine. Il est également possible, sur la base d'outils en pierre vieux de 2,1 millions d'années de Shangchen, en Chine, qu'une espèce ancestrale d'Homo rudolfensis et d'Homo habilis soit dispersée à travers l'Asie. Les estimations de la taille corporelle d'Homo rudolfensis et d'Homo habilis concluent généralement à une petite taille comparable à celle des australopithèques. Ces estimations dépendent en grande partie d'un seul squelette partiel d'Homo habilis qui a généré une estimation de poids comprise entre 44 et 82 livres.
On pense que Homo rudolfensis était plus grand que Homo habilis. Cependant, on ne sait pas exactement quelle était la taille de cette espèce. Aucun élément corporel n'a été définitivement associé à un crâne. Sur la base d'un crâne presque complet récupéré, on estime que le mâle Homo rudolfensis mesurait environ 5 pieds 3 pouces de hauteur et pesait environ 130 livres. On estime que les femelles mesuraient environ 4 pieds 11 pouces et pesaient 112 livres.
On suppose généralement que les hominidés pré-Homo ergaster présentaient un dimorphisme sexuel, les mâles étant nettement plus gros que les femelles. Cela comprendrait Homo rudolfensis et Homo habilis. Cependant, la masse corporelle relative des femelles est inconnue chez les deux espèces. On pense que les premiers hominidés avaient une épaisse couverture de poils sur le corps, comme les singes non humains modernes. En effet, ils semblent avoir habité des régions plus froides et auraient eu un mode de vie moins actif que les espèces post-ergaster présumées glabres. Ils auraient probablement eu besoin de poils épais pour rester au chaud. Cette évaluation inclut Homo rudolfensis.
Un spécimen juvénile d'Homo rudolfensis préservant un visage partiel a les mêmes repères d'âge qu'un humain moderne de 13 à 14 ans. Cependant, le juvénile Homo rudolfensis est probablement mort vers l'âge de 8 ans. Ceci est calculé sur un taux de croissance présumé plus rapide parmi les premiers hominidés, qui à son tour est basé sur le taux de développement dentaire. En 1983, des anthropologues examinant la base du crâne d'un spécimen d'Homo rudolfensis ont calculé un volume cérébral de 752 à 753 cc. En comparaison, les spécimens d'Homo habilis ont en moyenne environ 600 cc de volume cérébral. Homo ergaster avait en moyenne 850 cc de volume cérébral. Des études anthropologiques ultérieures ont estimé le volume cérébral d'Homo rudolfensis à 526 cc. Une étude encore plus tardive, réalisée en 2008, a estimé un volume cérébral estimé à 700 cc, basé sur le plus ancien spécimen connu d'Homo rudolfensis.
Les fossiles ont généralement été classés dans Homo rudolfensis en raison de la grande taille du crâne, du visage plus plat et plus large, des dents des joues plus larges, des couronnes et racines dentaires plus complexes et de l'émail plus épais que celui de l'Homo habilis. Les premiers Homo se caractérisent par des dents plus grandes que les Homo ultérieurs. On pense généralement que le régime alimentaire des premiers Homo comportait une plus grande proportion de viande que celui des Australopithèques. On suppose que cela a à son tour conduit à la croissance du cerveau. Les principales hypothèses à ce sujet consistent en la proposition selon laquelle la viande est plus riche en énergie et en nutriments que les autres sources alimentaires. Cela a exercé une pression évolutive sur le développement de compétences cognitives améliorées pour faciliter la récupération stratégique et monopoliser les carcasses fraîches. La viande a permis à l’intestin du singe, gros et coûteux en calories, de diminuer en taille, permettant ainsi à cette énergie d’être détournée vers la croissance du cerveau.
Alternativement, il est également suggéré que le début d’Homo se trouvait au milieu d’un climat sec avec des options alimentaires plus rares. Les premiers homos comptaient principalement sur les organes de stockage souterrains des légumes tels que les tubercules et le partage de nourriture. Cela a facilité les liens sociaux entre les membres masculins et féminins du groupe. Cependant, contrairement à ce qui est présumé pour Homo ergaster et plus tard Homo, les premiers Homo de petite taille étaient probablement incapables de courir et de chasser d'endurance. L'avant-bras long et semblable à un australopithèque d'Homo habilis pourrait indiquer que les premiers Homo étaient encore arboricoles dans une certaine mesure. On pense qu’une chasse et une cueillette coopératives bien organisées ont émergé chez Homo ergaster.
Cependant, les modèles de collecte de nourriture proposés pour expliquer la croissance importante du cerveau dicteraient nécessairement une augmentation de la distance de déplacement quotidienne. La grande taille des incisives chez Homo rudolfensis et Homo habilis par rapport aux prédécesseurs des Australopithecus implique que ces deux espèces s'appuient davantage sur les incisives. De grandes molaires ressemblant à des australopithèques pourraient indiquer une nourriture plus difficile sur le plan mécanique par rapport à Homo plus récent. Les corps des mandibules d'Homo rudolfensis et d'autres Homo anciens sont plus épais que ceux des humains modernes et de tous les singes vivants. Ils sont plus comparables aux australopithèques. Le corps mandibulaire résiste à la torsion due à la force de morsure ou à la mastication. Cela signifie que leurs mâchoires peuvent produire une pression inhabituellement puissante en mangeant.
Homo rudolfensis n’est associé à aucun outil. Cependant, le plus grand relief des cuspides molaires chez Homo rudolfensis et Homo habilis par rapport à l'Australopithèque suggère que les deux premiers utilisaient des outils pour fracturer des aliments durs tels que des parties de plantes souples ou de la viande. N'était-ce pas le cas, s'ils n'avaient pas utilisé d'outils pour transformer les aliments, les cuspides auraient été encore plus usées. Néanmoins, les adaptations de la mâchoire pour le traitement d’aliments mécaniquement difficiles indiquent que les progrès technologiques n’ont pas eu d’effet profond sur leur régime alimentaire.
De grandes concentrations d'outils en pierre sont connues sur Koobi Fora. Parce que ces agrégations coïncident avec l’émergence d’Homo ergaster, il est probable qu’Homo ergaster les ait fabriqués. Cependant, il n'est pas possible d'attribuer définitivement les outils à une espèce car Homo rudolfensis, Homo habilis et Paranthropus boisei sont également bien connus dans la région. Le spécimen le plus ancien d'Homo rudolfensis est associé à l'industrie des outils en pierre d'Oldowan, ce qui signifie que cette tradition était utilisée par le genre depuis son émergence.
Les premiers Homo rudolfensis et Paranthropus (à la fois Robustus et Bosei) ont des molaires exceptionnellement épaisses pour les hominidés. L’émergence de ces deux phénomènes coïncide avec une tendance au refroidissement et à l’aridité en Afrique il y a environ 2,5 millions d’années. Cela pourrait signifier qu’ils ont évolué en raison du changement climatique. Néanmoins, en Afrique de l’Est, les forêts tropicales et les zones boisées ont persisté malgré les périodes de sécheresse. Homo rudolfensis coexistait avec Homo habilis, Homo ergaster et Paranthropus boisei [Wikipedia].
Homo ergaster: Homo ergaster est une espèce ou sous-espèce éteinte d'humains archaïques qui vivaient en Afrique au début du Pléistocène. La question de savoir si Homo ergaster constitue une espèce à part entière ou s'il doit être intégré à Homo erectus est un débat permanent et non résolu au sein de la paléoanthropologie. Les partisans de la synonymisation désignent généralement Homo ergaster comme « Homo erectus africain » ou « Homo erectus ergaster ». Le nom Homo ergaster se traduit approximativement par « homme qui travaille ». Cela fait référence aux outils plus avancés utilisés par l’espèce par rapport à ceux de leurs ancêtres.
La gamme fossile d'Homo ergaster principalement couvre la période d'il y a 1,8 à 1,7 million d'années, avec une poignée de spécimens plus âgés et plus jeunes étendant la gamme à il y a environ 2 et 1,5 millions d'années respectivement. Bien que des fossiles soient connus dans toute l’Afrique de l’Est et australe, la plupart des fossiles d’Homo ergaster ont été découverts le long des rives du lac Turkana au Kenya. Il existe des fossiles africains plus récents, certains datant de moins d'un million d'années, qui indiquent une continuité anatomique à long terme. Cependant, il n'est pas certain qu'ils puissent être correctement considérés comme des spécimens d'Homo ergaster. En tant que chrono-espèce, Homo ergaster a peut-être persisté il y a 600 000 ans, lorsque de nouvelles lignées d'Homo sont apparues en Afrique.
Les chercheurs qui pensent qu'Homo ergaster devrait être intégré à Homo erectus considèrent qu'il y a trop peu de différence entre les deux pour les séparer en espèces distinctes. Les partisans du maintien des deux espèces distinctes citent des différences morphologiques entre les fossiles africains et les fossiles d'Homo erectus d'Asie. Ils proposent également que l'évolution précoce d'Homo était bien plus complexe que ce que laisse entendre l'intégration d'espèces telles que Homo ergaster dans Homo erectus. De plus, les différences morphologiques entre les spécimens communément considérés comme constituant Homo ergaster pourraient suggérer que Homo ergaster lui-même ne représente pas une espèce cohésive. Il peut y avoir plus d'une espèce distincte identifiée à tort comme Homo ergaster.
Quelle que soit leur classification la plus correcte, Homo ergaster présente des versions primitives de traits exprimés plus tard chez Homo erectus. Cela amène les chercheurs à croire que l'Homo ergaster est probablement l'ancêtre direct des populations ultérieures d'Homo erectus en Asie. De plus, Homo ergaster est probablement l'ancêtre des hominidés ultérieurs d'Europe et d'Afrique, tels que les humains modernes (Homo sapien) et les Néandertaliens. Plusieurs caractéristiques distinguent Homo ergaster des australopithèques ainsi que des espèces d'Homo plus anciennes et plus basales, comme Homo habilis. Parmi ces caractéristiques figurent leur masse corporelle plus importante, leurs jambes relativement longues et leur bipédie obligatoire. Des caractéristiques telles que des mâchoires et des dents relativement petites sont également distinctives, ce qui indique un changement majeur dans le régime alimentaire. Des caractéristiques distinctives supplémentaires incluent les proportions du corps.
La dernière et conséquence de ces distinctions physiologiques serait les modes de vie déduits que les chercheurs présument être plus similaires à ceux des humains modernes qu'à ceux des hominidés antérieurs et contemporains. Compte tenu de ces caractéristiques, certains chercheurs considèrent Homo ergaster comme étant le premier véritable représentant du genre Homo. Homo ergaster vivait dans la savane en Afrique. Il s’agissait d’un environnement unique avec des défis qui auraient entraîné la nécessité d’adopter de nombreux comportements nouveaux et distincts. Auparavant, Homo utilisait probablement des tactiques de contre-attaque, comme les primates modernes, pour éloigner les prédateurs. À l’époque d’Homo ergaster, ce comportement avait probablement abouti au développement d’un véritable comportement de chasseur-cueilleur, une première chez les primates.
D'autres comportements qui auraient pu apparaître pour la première fois chez Homo ergaster incluent les divisions mâles-femelles de recherche de nourriture et de véritables liens de couple monogames. Homo ergaster marque également l'apparition d'outils plus avancés de l'industrie acheuléenne, notamment les premières haches à main connues. Bien que des preuves incontestées manquent, Homo ergaster pourrait également avoir été le premier hominidé à maîtriser le contrôle du feu. La systématique et la taxonomie d'Homo du Pléistocène inférieur au Pléistocène moyen sont l'un des domaines les plus controversés de la paléoanthropologie. Au début de la paléoanthropologie et jusqu'au XXe siècle, il était généralement supposé que H. sapiens était le résultat final de modifications progressives au sein d'une seule lignée d'évolution d'hominidés.
L'Homo erectus était perçu comme la forme de transition entre les premiers hominidés et les humains modernes. La vaste classification d’Homo erectus a été initialement « érigée » pour contenir des fossiles humains archaïques en Asie. La classification en est venue à englober une gamme extraordinairement large de fossiles couvrant une période de temps énorme. En fait, la période initialement attribuée à Homo erectus était presque toute la plage temporelle d'Homo. Depuis la fin du XXe siècle, l’énorme diversité au sein d’Homo erectus a conduit certains chercheurs à se demander ce qui définit exactement l’espèce et ce qu’elle devrait englober. Certains chercheurs comme le paléoanthropologue Ian Tattersall en 2013 ont remis en question le caractère trop large de la classification Homo erectus. Selon leur position, il contient simplement un nombre « lourd » de fossiles avec des « morphologies sensiblement différentes ».
Dans les années 1970, les paléoanthropologues Richard Leakey et Alan Walker ont décrit une série de fossiles d'hominidés provenant de la rive est du lac Turkana au Kenya. Les découvertes les plus notables étaient deux crânes partiels trouvés à Koobi Fora. Leakey et Walker ont attribué ces crânes à Homo erectus, notant que leurs volumes cérébraux (848 et 803 cc respectivement) se comparaient bien au spécimen type beaucoup plus jeune d'Homo erectus (950 cc). Un autre fossile important était une mandibule fossile récupérée à Ileret et décrite par Leakey en 1972 comme « Homo d'espèce indéterminée ».
En 1975, les paléoanthropologues ont désigné les fossiles comme spécimen holotype d'une espèce distincte, qu'ils ont surnommée Homo ergaster. Le nom ergaster est dérivé du grec ancien et se traduit approximativement par « ouvrier » ou « ouvrier ». Un fossile presque complet a été découvert sur la rive ouest du lac Turkana en 1984 par des archéologues kenyans. Interprétés comme étant le squelette d'un jeune mâle (bien que le sexe soit en réalité indéterminé), les fossiles ont été décrits par Leakey et Walker et surnommés « Turkana Boy ». Le fossile consistait en un squelette presque complet. Il a été décrit par Leakey et Walker comme représentant l'Homo erectus.
« Turkana Boy » a été le premier spécimen entièrement préservé d'Homo ergaster/erectus découvert et constitue un fossile important pour établir les différences et les similitudes entre les premiers Homo et les humains modernes. Turkana Boy a finalement été classé comme Homo ergaster en 1992. Aux côtés d’autres fossiles d’Afrique précédemment/à tort classés comme Homo erectus, « Turkana Boy » est communément considéré comme un représentant d’Homo ergaster par ceux qui soutiennent Homo ergaster comme une espèce distincte.
Homo ergaster se distingue facilement des espèces antérieures et plus basales d'Homo, notamment Homo habilis et Homo rudolfensis. Il existe un certain nombre de caractéristiques qui les alignent, ainsi que leur mode de vie déduit, plus étroitement sur les humains modernes que sur les hominidés antérieurs et contemporains. Par rapport à leurs parents, Homo ergaster avait des proportions corporelles plus similaires à celles des membres ultérieurs du genre Homo. Celles-ci incluraient notamment des jambes relativement longues qui les auraient rendus obligatoirement bipèdes. Les dents et les mâchoires d'Homo ergaster sont également relativement plus petites que celles d'Homo habilis et d'Homo rudolfensis. Cela indique un changement majeur dans le régime alimentaire et, par conséquent, dans le mode de vie.
En 1999, des paléoanthropologues ont affirmé que les critères conventionnels d'attribution des espèces au genre Homo étaient erronés. Ils ont proposé que les espèces précoces et basales telles que Homo habilis et Homo rudolfensis pourraient être reclassées de manière appropriée comme australopithèques ancestraux. Selon eux, le véritable premier représentant d'Homo était Homo ergaster. Depuis sa description en tant qu'espèce distincte en 1975, la classification des fossiles référencés à Homo ergaster est controversée. Homo ergaster a été immédiatement rejeté par Leakey et Walker ainsi que par de nombreux chercheurs influents. En 1990, les partisans de ce point de vue à contre-courant ont écrit un traité plaidant en faveur d’une définition plus inclusive et plus large de l’Homo erectus.
Dans l’ensemble, il ne fait aucun doute que le groupe de fossiles composant Homo erectus et Homo ergaster représente les fossiles d’un sous-ensemble plus ou moins cohérent d’humains archaïques étroitement liés. La question est plutôt de savoir si ces fossiles représentent un rayonnement d’espèces différentes ou le rayonnement d’une espèce unique, très variable et diversifiée, sur une période de près de deux millions d’années. Ce débat de longue date n’est toujours pas résolu. Aujourd'hui, les chercheurs utilisent généralement le terme Homo erectus ss (sensu stricto) pour désigner les fossiles d'Homo erectus en Asie. Ils utilisent le terme Homo erectus sl (sensu lato) pour désigner les fossiles d'autres espèces qui peuvent ou non être inclus dans Homo erectus. . Cette terminologie comprendrait Homo ergaster, Homo antecessor et Homo. heidelbergensis.
Pour des raisons évidentes, Homo ergaster partage de nombreuses caractéristiques avec Homo erectus. Ces caractéristiques communes comprennent de grandes mâchoires projetées vers l'avant, de grandes arcades sourcilières et un front fuyant. De nombreuses caractéristiques de l'Homo ergaster sont clairement des versions plus primitives de caractéristiques exprimées plus tard chez l'Homo erectus. Cela obscurcit quelque peu les différences entre les deux. Il existe cependant des différences subtiles, potentiellement significatives, entre les fossiles d’Afrique de l’Est et d’Asie de l’Est. Parmi ceux-ci se trouvent les crânes d'Homo ergaster, un peu plus hauts et aux parois plus fines. Également les arcades sourcilières et les visages encore plus massifs de l'Homo erectus asiatique.
La question est d'autant plus difficile qu'elle concerne le degré de variation intraspécifique qui peut être manifesté chez une seule espèce avant que cette variation nécessite la définition d'une espèce distincte. Cette question en soi n’a pas de réponse claire. Lors d'une analyse de 2008, un examen des fossiles de diverses sous-espèces d'Homo erectus, y compris des fossiles attribués à Homo ergaster, a été réalisé. L'étude a conclu que la variation intraspécifique au sein de l'Homo erectus était plus grande que prévu pour une seule espèce par rapport aux humains et aux chimpanzés modernes. Cependant, l'étude a également révélé que la variation intraspécifique correspondait bien à la variation attendue pour une espèce par rapport aux gorilles. Il se situe même bien dans la fourchette attendue pour une seule sous-espèce par rapport aux orangs-outans. Ce dernier phénomène était cependant en partie dû au grand dimorphisme sexuel présenté chez les gorilles et les orangs-outans.
L'étude a conclu que Homo erectus sl était soit une espèce unique mais variable, soit plusieurs sous-espèces divisées par le temps et la géographie, soit plusieurs espèces géographiquement dispersées mais étroitement liées. Une étude de 2015 a répertorié Homo ergaster comme l'une des sept espèces d'Homo « largement reconnues », aux côtés de Homo habilis, Homo rudolfensis, Homo erectus, Homo heidelbergensis, Homo neanderthalensis et Homo sapiens. L'étude a également noté que plusieurs autres espèces telles que Homo floresiensis et Homo antecessor étaient moins largement reconnues ou plus mal connues.
Une autre étude de 2013 a comparé divers fossiles africains attribués à Homo erectus ou Homo ergaster à des fossiles asiatiques, notamment le spécimen type d'Homo erectus. La conclusion de l'étude était que faire référence au matériel africain comme Homo ergaster plutôt que "Homo erectus africain" était une "amélioration considérable" car il existait de nombreuses autapomorphies distinguant le matériel des deux continents l'un de l'autre. L'étude a proposé qu'il serait approprié d'utiliser la désignation Homo erectus uniquement pour les fossiles d'Asie de l'Est. Que son utilisation antérieure comme nom pour une qualité adaptative de fossiles humains provenant de toute l’Afrique et de l’Eurasie devrait être abandonnée.
Bien que l'on suppose souvent qu'elle est originaire d'Afrique de l'Est, les origines d'Homo ergaster sont obscurcies par le fait que l'espèce marque un changement radical par rapport aux espèces antérieures d'Homo et d'Australopithèque par ses membres longs, sa hauteur et ses proportions corporelles modernes. Bien qu'un grand nombre d'outils du Pléistocène aient été découverts en Afrique de l'Est, on ne peut pas être entièrement sûr de l'origine de l'Homo ergaster sans d'autres découvertes de fossiles. On suppose qu'Homo ergaster a évolué à partir d'espèces antérieures d'Homo, probablement Homo habilis.
Les populations d'Homo ergaster en dehors de l'Afrique ont été déduites sur la base de la répartition géographique de leurs descendants et d'outils correspondant à ceux d'Afrique de l'Est. Cependant, les fossiles de l’espèce proviennent principalement d’Afrique de l’Est, il y a 1,8 à 1,7 millions d’années. La plupart des fossiles ont été récupérés sur les rives du lac Turkana au Kenya. Le plus ancien spécimen connu d'Homo ergaster est un crâne découvert dans le système paléocave de Drimolen en Afrique du Sud. Il est daté d’il y a plus de 1,95 millions d’années. Cela démontre que les premiers Homo ergaster coexistaient avec d'autres hominidés tels que Paranthropus Robustus et Australopithecus sediba.
Il existe également des spécimens plus jeunes d'Homo ergaster. Plus particulièrement, Turkana Boy est daté d'il y a environ 1,56 million d'années. Une poignée de crânes africains encore plus jeunes plaident en faveur d’une continuité anatomique à long terme. Cependant, il n'est pas certain qu'ils puissent être formellement considérés comme des spécimens d'Homo ergaster. Il y a un crâne des gorges d'Olduvai connu sous le nom de "Olduvai Hominid 9", daté d'il y a environ 1,2 à 1,1 million d'années. Il existe également des crânes de Buia (près de la côte érythréenne) datés d'il y a environ 1 million d'années. Il existe des crânes encore plus jeunes provenant d'Homo ergaster provisoirement identifiés dans la formation de Bouri en Éthiopie, datés entre 1 million et 780 000 ans. Enfin, il y a un crâne fragmentaire d'Olorgesailie au Kenya daté d'il y a entre 970 000 et 900 000 ans.
Le crâne d'Olduvai est similaire à l'Homo erectus asiatique dans sa crête sourcilière massive, mais les autres ne présentent que des différences mineures par rapport aux crânes antérieurs d'Homo ergaster. L'Homo erectus en Asie ainsi que les hominidés ultérieurs en Europe et en Afrique sont tous probablement des lignées descendantes de l'Homo ergaster. Cela comprendrait l'Homo heidelbergensis et l'Homo neanderthalensis d'Europe, ainsi que l'Homo sapiens d'Afrique. Étant donné que l'on pense qu'Homo ergaster était l'ancêtre de ces Homo ultérieurs, il pourrait avoir persisté en Afrique jusqu'à il y a environ 600 000 ans. Ce point s’étendrait théoriquement jusqu’au moment où la taille de son cerveau a augmenté rapidement et où Homo heidelbergensis a émergé.
Traditionnellement, l’Homo erectus était considéré comme l’hominidé qui avait quitté l’Afrique pour la première fois pour coloniser l’Europe et l’Asie. Si Homo ergaster est distinct de Homo erectus, ce rôle s'appliquerait plutôt à Homo ergaster. Très peu d'informations concrètes sont connues sur quand et quels Homo sont apparus pour la première fois en Europe et en Asie, car les hominines fossiles du Pléistocène inférieur sont rares sur les deux continents. La question de savoir si ce serait Homo ergaster (ou « les premiers Homo erectus ») qui s'est étendu en Europe et en Asie, ainsi que la manière particulière dont ils l'ont fait, reste une conjecture.
La présence de fossiles d'Homo erectus en Asie de l'Est signifie qu'une espèce humaine, très probablement Homo ergaster, avait quitté l'Afrique il y a moins d'un million d'années. Historiquement, l’hypothèse était qu’ils avaient migré pour la première fois hors d’Afrique il y a environ 1,9 à 1,7 millions d’années. Les découvertes en Géorgie et en Chine repoussent la dernière date possible plus loin, avant 2 millions d'années. Cependant, cela remet également en question l’idée selon laquelle Homo ergaster aurait été le premier hominidé à quitter l’Afrique.
La principale raison de quitter l’Afrique est probablement une population croissante qui dépasse périodiquement sa base de ressources. Des groupes dissidents se seraient déplacés pour s’établir dans les territoires vides voisins au fil du temps. La physiologie et la technologie améliorée d’Homo ergaster auraient pu leur permettre de voyager et de coloniser des territoires que personne n’avait jamais occupés auparavant. On ne sait pas vraiment si Homo ergaster était réellement capable de s’étendre en dehors de l’Afrique. Les australopithèques avaient probablement colonisé les prairies de savane dans toute l'Afrique il y a 3 millions d'années. Il n’y a aucune raison claire pour laquelle ils n’auraient pas pu s’étendre dans les prairies d’Asie avant Homo ergaster.
L’hypothèse générale est que les hominidés ont migré hors du continent soit par l’extrémité sud de la mer Rouge, soit le long de la vallée du Nil. Cependant, il n'existe aucun hominidé fossile connu dans l'une ou l'autre région du Pléistocène inférieur. Les premiers fossiles d'Homo en dehors de l'Afrique sont les crânes de Dmanisi de Géorgie. Ceux-ci sont datés de 1,77 à 1,85 millions d’années. Ils représentent soit le premier Homo ergaster, soit un nouveau taxon, Homo georgicus. Ensuite, il y avait trois incisives d'Ubeidiya en Israël, datées d'environ 1,4 à 1 million d'années. Et bien sûr, il y avait les célèbres fossiles de l’homme de Java, Homo erectus, découverts à plus de huit mille kilomètres de là.
La datation des principaux spécimens asiatiques d’Homo erectus n’est pas entièrement certaine, mais ils sont probablement tous âgés de 1,5 million d’années ou moins. Cela inclut Java Man. Ubeidiya, en Israël, est également le plus ancien site fermement confirmé d'outils acheuléens en dehors de l'Afrique. Les outils récupérés là-bas ressemblent beaucoup à des outils plus anciens découverts en Afrique de l'Est. Les outils acheuléens sont l'une des industries d'outils associées à Homo ergaster. Les premières preuves fossiles d'Homo en Asie sont les crânes de Dmanisi de Géorgie. Ceux-ci partagent de nombreux traits avec Homo ergaster en Afrique. Cela suggère qu’Homo ergaster pourrait s’être répandu hors d’Afrique il y a 1,7 à 1,9 millions d’années.
En plus des traits de type Homo ergaster, les crânes de Dmanisi possèdent un large assortiment d'autres traits. Certains de ces traits sont similaires à ceux des hominidés antérieurs tels que Homo habilis. Cependant, le site manque notamment et peut-être de manière significative de haches à main préservées. Les haches à main sont par ailleurs caractéristiques de l'Homo ergaster. Cela implique que les hominidés pourraient s'être répandus hors d'Afrique encore plus tôt qu'Homo ergaster. L'un des crânes de Dmanisi ressemble particulièrement à Homo habilis par le petit volume de son coffret crânien (environ 600 cc). La forme du visage moyen et supérieur et l'absence de nez externe rappellent également Homo habilis.
Le mélange de crânes à Dmanisi suggère que la définition de Homo ergaster (ou Homo erectus) pourrait être élargie de manière plus appropriée. Peut-être que la définition devrait s'étendre aux fossiles qui autrement seraient attribués à Homo habilis. Ou peut-être devrait-il reconnaître que deux espèces distinctes d’humains archaïques ont quitté l’Afrique très tôt. Outre les fossiles de Dmanisi, des outils en pierre fabriqués par des hominidés ont été découverts sur le plateau de Loess en Chine. Ces outils étaient datés de 2,12 millions d’années. C’est une preuve claire que les hominidés ont dû quitter l’Afrique avant cette époque.
Historiquement, une hypothèse alternative était qu'Homo aurait évolué en Asie à partir d'ancêtres antérieurs qui y avaient émigré depuis l'Afrique. Par la suite, cet ancêtre antérieur s’est répandu en Europe, où il a donné naissance à Homo sapiens. Ce point de vue était notamment partagé par les anthropologues du XIXe et du début du XXe siècle, dont Eugène Dubois. Dubois a décrit pour la première fois les fossiles d'Homo erectus au 19ème siècle et a examiné les fossiles de l'Homme de Java. À l’époque, les fossiles de l’Homme de Java étaient indéniablement les premiers fossiles d’hominidés connus. Ces fossiles ont été proposés comme preuve de l'hypothèse d'une migration des premiers hominidés hors d'Asie et vers l'Europe.
La découverte des australopithèques et d'Homo antérieurs en Afrique signifiait qu'Homo lui-même n'était pas originaire d'Asie. Cependant, l'idée a parfois refait surface selon laquelle l'Homo erectus (ou Homo ergaster) en particulier était originaire d'Asie et s'est ensuite répandu en Afrique. Diverses découvertes de fossiles ont été utilisées pour étayer cette proposition au fil des ans. L’exemple le plus célèbre est peut-être un ensemble massif de mâchoires provenant d’Indonésie. Ils étaient perçus comme semblables à ceux des australopithèques et surnommés Meganthropus. Les anthropologues pensent maintenant que les fossiles appartiennent à un singe hominidé sans lien de parenté.
La découverte d'Homo floresiensis en 2003 a de nouveau conduit à des suggestions d'hominines pré-erectus en Asie. Homo floresiensis a conservé une anatomie primitive du pied et du poignet qui rappelle celle de Homo habilis et de l'Australopithèque. Cependant, il n’existe aucun os comparable du pied ou du poignet d’Homo erectus, ce qui rend les comparaisons directes impossibles. L'idée selon laquelle Homo ergaster/Homo erectus a d'abord évolué en Asie avant de s'étendre à nouveau en Afrique a été considérablement affaiblie par la datation du crâne trouvé dans le système paléocave de Drimolen en Afrique du Sud, âgé d'environ 2 millions d'années. Cette découverte est antérieure à tous les autres fossiles connus d’Homo ergaster/Homo erectus.
Les seuls restes post-crâniens bien conservés d'Homo ergaster proviennent du fossile de Turkana Boy. Contrairement aux australopithèques, les bras de Turkana Boy n'étaient pas plus longs par rapport à ses jambes que les bras des humains modernes. Le torse en forme de cône des ancêtres de Turkana Boy avait évolué vers une poitrine plus en forme de tonneau sur des hanches étroites. C'est une autre similitude avec les humains modernes. Le tibia (os du tibia) du Turkana Boy est relativement plus long que le même os chez l'homme moderne, ce qui signifie potentiellement que le genou était plus plié lors de la marche.
La silhouette mince et longue du Turkana Boy peut s'expliquer par le fait qu'Homo ergaster vit dans des environnements saisonniers chauds et arides. En raison de l’amincissement du corps, le volume corporel diminue plus rapidement que la surface cutanée et une plus grande surface cutanée par rapport au volume corporel signifie une dissipation thermique plus efficace. Les individus Homo ergaster étaient nettement plus grands que leurs ancêtres. Lucy, le célèbre fossile d'australopithèque, ne mesurait qu'environ 1 mètre (3 pieds, 3 pouces) à sa mort. Turkana Boy mesurait environ 1,62 mètre (5 pieds 4 pouces) lorsqu'il était juvénile. Turkana Boy aurait probablement atteint 1,82 mètre ou plus s'il avait survécu jusqu'à l'âge adulte. On pense que l'Homo ergaster adulte mesurait entre 1,45 et 1,85 mètre (4 pieds 9 pouces à 6 pieds 1 pouce).
En raison de son adaptation à un climat chaud et aride, Homo ergaster pourrait également avoir été la première espèce humaine à avoir une peau presque glabre et nue. La transpiration est le principal moyen par lequel les humains modernes empêchent leur cerveau et leur corps de surchauffer. Si Homo ergaster avait eu une couverture de poils semblable à celle d'un singe, la transpiration n'aurait pas été aussi efficace. Bien que la transpiration soit l'explication généralement acceptée de l'absence de poils, d'autres explications proposées incluent une réduction de la charge parasitaire et de la sélection sexuelle.
Il est douteux que les australopithèques et les premiers Homo soient suffisamment mobiles pour faire de la perte de cheveux un trait avantageux. Cependant, Homo ergaster était clairement adapté aux voyages sur de longues distances et connu pour habiter à des altitudes plus basses que ses ancêtres. Les australopithèques habitaient généralement des altitudes plus froides et plus élevées, de 3 300 à 5 200 pieds. Les températures nocturnes seraient devenues beaucoup plus froides et il aurait peut-être fallu isoler les poils du corps. Homo ergaster, en revanche, se trouvait principalement dans des environnements de savane ouverts et chauds.
Alternativement et malgré cela, la perte de poils aurait pu survenir beaucoup plus tôt que Homo ergaster. Bien que les impressions cutanées soient inconnues chez les hominidés éteints, il est possible que les ancêtres humains perdaient déjà leurs poils il y a environ 3 millions d'années. Les ancêtres humains ont contracté les poux du pubis des gorilles il y a environ 3 millions d'années. La spéciation des poux du pubis entre les humains et les gorilles n'était potentiellement possible que parce que les ancêtres humains avaient perdu la plupart de leurs poils à cette date précoce. Il est également possible que la perte de poils se soit produite à une date beaucoup plus tardive.
L'analyse génétique suggère une activité élevée du récepteur de la mélanocortine 1 qui produit une peau foncée. Cette activité génétique remonte à environ 1,2 million d'années. Cela pourrait indiquer l’évolution de la calvitie à cette époque. Un manque de poils avant cette période aurait exposé la peau à des rayons UV nocifs. Les différences avec les humains modernes auraient été facilement apparentes sur le visage et le crâne d'Homo ergaster. Le cerveau de Turkana Boy était presque entièrement développé au moment de sa mort. Cependant, son volume à 880 cc n'était qu'environ 130 cc supérieur au maximum trouvé chez Homo habilis. Et son volume cérébral était environ 500 cc inférieur à la moyenne des humains modernes.
L'augmentation de 130 cc de la taille du cerveau par rapport à Homo habilis devient beaucoup moins significative que ce que l'on pourrait présumer lorsque l'on considère la plus grande taille corporelle de Turkana Boy et d'Homo ergaster. Pour tous les crânes d'Homo ergaster considérés, le volume cérébral de l'espèce variait généralement entre 600 et 910 cc. Certains exemplaires plus petits n'avaient qu'un volume de 508 à 580 cm3. Comme leur cerveau était plus petit que celui des humains modernes, le crâne d’Homo ergaster s’est immédiatement rétréci derrière les orbites (constriction post-orbitale).
Le cas cérébral de Turkana Boy était long et bas. Son front aurait été plat et fuyant, fusionnant selon un angle avec l'arcade sourcilière au-dessus de ses yeux. Une différence notable entre Turkana Boy et les australopithèques et Homo habilis aurait été son nez. Le nez de Turkana Boy aurait été similaire à celui des humains modernes en se projetant vers l'avant et en ayant les narines orientées vers le bas. Ce nez externe pourrait aussi être une adaptation vers un climat plus chaud. Le nez des humains modernes est généralement plus froid que celui de leur corps central. Cela condense l'humidité qui aurait autrement été expirée et perdue pendant les périodes d'activité accrue.
Le visage de Turkana Boy aurait été plus long de haut en bas que celui des humains modernes, avec les mâchoires projetées plus loin vers l'extérieur (prognathisme). Même si les mâchoires et les dents étaient plus petites que celles de l'australopithèque moyen et de l'Homo habilis, elles étaient néanmoins nettement plus grandes que celles de l'homme moderne. Comme la mâchoire était fortement inclinée vers l'arrière, il est probable qu'il n'avait pas de menton. La structure globale du crâne et du visage de Turkana Boy se reflète également dans d'autres crânes d'Homo ergaster. Ceux-ci combinent des visages larges et saillants vers l’extérieur avec des arcades sourcilières, des fronts fuyants, de grandes dents et des os nasaux saillants.
Turkana Boy n'aurait pas plus de 12 ans lorsqu'il est mort. Sa stature ressemble davantage à celle d’un jeune de 15 ans moderne. Le cerveau est comparable à celui d’un enfant moderne d’un an. Selon les normes modernes, Homo ergaster aurait donc été limité sur le plan cognitif. Néanmoins, l’invention de nouveaux outils prouve qu’ils étaient plus intelligents que leurs prédécesseurs. Homo ergaster possédait également une masse corporelle nettement plus importante que les hominidés antérieurs tels que les premiers Homo, Australopithecus et Paranthropus.
Les australopithèques pesaient généralement entre 64 et 106 livres. À titre de comparaison, le poids d'Homo ergaster variait généralement entre 115 et 139 livres. Il est possible que l’augmentation de la taille du corps soit le résultat de la vie dans un environnement de savane ouverte. Une taille accrue dans cet environnement leur donnerait la possibilité d’exploiter des régimes alimentaires plus larges dans des zones d’alimentation plus vastes. Une plus grande taille augmenterait également la mobilité et améliorerait la capacité de chasser des proies plus grosses. L'augmentation de la masse corporelle signifie également que les parents auraient pu porter leurs enfants à un âge plus avancé et avec une masse plus importante.
Le dimorphisme sexuel réduit (différences entre la taille des mâles et des femelles, etc.) a souvent été cité historiquement comme l'une des différences radicales entre Homo ergaster et les Homo et australopithèques antérieurs. Cependant, en réalité, il n'est pas clair si les australopithèques étaient significativement plus dimorphes sexuellement que l'Homo ergaster ou les humains modernes. Des preuves squelettiques suggèrent que les sexes chez Homo ergaster ne différaient pas plus en taille que ceux des humains modernes. En outre, une étude paléoanthropologique de 2003 a suggéré que la même chose était également vraie pour l'Australopithecus afarensis, beaucoup plus ancien.
Le dimorphisme sexuel est difficile à mesurer chez les espèces disparues puisque le sexe des fossiles n'est généralement pas déterminable. Historiquement, les scientifiques ont généralement mesuré les différences entre les extrémités (en termes de taille et de morphologie) du matériel fossile attribué à une espèce et ont supposé que le rapport obtenu s'appliquait à la différence moyenne entre les individus mâles et femelles. Mais cette généralisation simplifiée peut être exacte ou non.
Les dimensions du bassin d'une femme adulte Homo ergaster âgée de 1,8 million d'années, originaire de Gona, en Éthiopie, suggèrent qu'Homo ergaster aurait été capable de donner naissance à des enfants avec une taille cérébrale prénatale (avant la naissance) maximale de 315 cc. Cela représente environ 30 à 50 % de la taille du cerveau adulte. Cette valeur se situe entre celle des chimpanzés (environ 40 %) et celle de l'homme moderne (28 %). D'autres conclusions sur la croissance et le développement du jeune Homo peuvent être tirées de l'enfant Mojokerto. L'enfant Mojokerto est un Homo erectus asiatique âgé d'environ un an. Il est daté d'environ 1,4 à 1,5 millions d'années. Il possédait un cerveau d'environ 72 à 84 % de la taille d'un cerveau d'Homo erectus adulte. Cela suggère une trajectoire de croissance cérébrale plus similaire à celle des autres grands singes qu’à celle des humains modernes.
Le bassin Gona et l'enfant Mojokerto suggèrent que la croissance prénatale d'Homo ergaster était similaire à celle des humains modernes. Cependant, ils suggèrent également que la croissance et le développement postnatals (après la naissance) étaient intermédiaires entre ceux des chimpanzés et des humains modernes. Le taux de développement plus rapide suggère que l’altricialité a évolué à un stade ultérieur de l’évolution humaine. L'altricialité est une enfance prolongée et une longue période de dépendance parentale. L'altricalité chez Homo ne s'est peut-être pas développée jusqu'au dernier ancêtre commun des Néandertaliens et des humains modernes. Le taux de développement plus rapide pourrait également indiquer que la durée de vie attendue de l’Homo ergaster et de l’Homo erectus était inférieure à celle des humains plus récents et modernes.
Comparé à ses ancêtres, Homo ergaster possédait un corps et un cerveau plus grands. On suppose souvent que ces attributs auraient entraîné une augmentation des besoins alimentaires et énergétiques. En 2002, des paléoanthropologues ont calculé que les besoins métaboliques moyens au repos d'Homo ergaster auraient été 39 % plus élevés que ceux d'Australopithecus afarensis, 30 % plus élevés chez les mâles et 54 % plus élevés chez les femelles. Cependant, les proportions du torse d'Homo ergaster impliquent un intestin relativement petit. Cela pourrait signifier que les besoins énergétiques n’étaient pas nécessairement plus élevés chez Homo ergaster que chez les hominidés antérieurs.
Cela serait dû au fait que les intestins des premiers singes (et des australopithèques) étaient gros et coûteux en énergie, car ils devaient synthétiser la graisse et l'énergie par la fermentation de la matière végétale. Homo ergaster, en revanche, mangeait probablement beaucoup plus de graisse animale que ses prédécesseurs. Cela aurait permis de consacrer davantage d’énergie à la croissance du cerveau. Cela pourrait théoriquement avoir augmenté la taille du cerveau sans nécessiter des besoins énergétiques supérieurs à ceux des espèces antérieures. Si en fait leurs besoins énergétiques avaient augmenté, Homo ergaster aurait eu besoin de manger soit beaucoup plus de nourriture que les australopithèques, soit aurait eu besoin de manger des aliments à plus forte teneur énergétique.
Si Homo ergaster mangeait le même type d'aliments que les australopithèques, le temps d'alimentation aurait alors dû être considérablement augmenté proportionnellement aux calories supplémentaires nécessaires. Cela aurait réduit le temps qu'Homo ergaster pouvait consacrer au repos, à la socialisation et aux voyages. Même si cela aurait été possible, cela semble peu probable. Cela est d’autant plus vrai que les mâchoires et les dents d’Homo ergaster sont de taille réduite par rapport à celles des australopithèques. Cette caractéristique suggère un changement de régime alimentaire, abandonnant les aliments fibreux et difficiles à mâcher. Quels que soient les besoins énergétiques, le petit intestin d’Homo ergaster suggère également un régime alimentaire plus facile à digérer, composé d’aliments de densité calorique plus élevée.
Il est probable qu'Homo ergaster consommait de la viande dans des proportions plus élevées que les australopithèques antérieurs. La viande a probablement été obtenue grâce à une combinaison d'embuscades, de chasse active et de fouilles conflictuelles. Homo ergaster devait sûrement posséder la capacité de courir en endurance pour ne pas devenir un repas. Mais Homo ergaster doit également avoir été capable de se défendre ainsi que les carcasses de ses proies contre la variété de prédateurs et charognards africains contemporains. Il est même possible qu’une diminution de la diversité des espèces carnivores africaines il y a environ 1,5 million d’années puisse être attribuée à la compétition avec les hominines opportunistes et carnivores.
À elle seule, la viande n’aurait peut-être pas suffi à nourrir pleinement l’Homo ergaster. Les humains modernes ne peuvent pas métaboliser suffisamment les protéines pour répondre à plus de 50 % de leurs besoins énergétiques. Les humains modernes qui dépendent fortement de produits d’origine animale dans leur alimentation dépendent principalement des graisses pour subvenir au reste de leurs besoins énergétiques. Plusieurs raisons rendent improbable un régime entièrement à base de viande chez Homo ergaster. La plus importante de ces raisons serait que les ongulés africains (les mammifères ongulés, les principales proies disponibles) sont relativement pauvres en graisse. Les régimes riches en viande nécessitent une consommation accrue d’eau. Répondre à de tels besoins aurait été difficile dans un environnement ouvert et chaud.
Les chasseurs-cueilleurs africains modernes qui dépendent fortement de la viande utilisent des moyens culturels pour récupérer le maximum de graisse des carcasses de leurs proies. Les méthodes utilisées par des peuples tels que les Hadza et les San n'auraient pas été accessibles à Homo ergaster. Homo ergaster aurait probablement consommé de grandes quantités de viande, bien plus que leurs ancêtres. Cependant, ils auraient également dû utiliser diverses autres sources de nourriture, telles que des graines, du miel, des noix, des invertébrés, des tubercules nutritifs, des bulbes et d'autres organes de stockage souterrains de plantes.
De plus, Homo ergaster possédait une capacité de mastication relativement faible. Cela est particulièrement vrai par rapport à ses ancêtres aux mâchoires plus grandes. Cela signifie que la viande et les aliments végétaux consommés auraient probablement nécessité l’utilisation d’outils pour être transformés avant d’être consommés. Homo ergaster vivait dans la savane africaine. Au Pléistocène, cette région abritait une communauté de carnivores considérablement plus formidable qu'aujourd'hui. Les hominidés n'auraient probablement pu s'adapter à la vie dans la savane que si des comportements de défense efficaces contre les prédateurs avaient déjà évolué. La défense contre les prédateurs serait probablement venue de l'Homo ergaster vivant en grands groupes. De plus, ils devaient posséder des outils en pierre (et probablement en bois) qu'ils utilisaient pour contre-attaquer efficacement.
Chez les primates modernes qui passent beaucoup de temps dans la savane, les individus forment de grands groupes composés de plusieurs mâles. Cela inclurait les chimpanzés contemporains et les babouins de la savane. Plusieurs mâles peuvent travailler efficacement ensemble pour repousser et contre-attaquer les prédateurs. Parfois même des pierres ou des bâtons sont employés pour défendre le reste du groupe. Il est possible qu'un comportement similaire ait été présenté au début d'Homo. Sur la base des systèmes liés aux mâles chez les bonobos et les chimpanzés, les groupes des premiers Homo pourraient également avoir été liés aux mâles. Cette possibilité est renforcée par la tendance à la création de liens masculins chez les primates butineurs modernes.
En raison de la rareté des matériaux fossiles, la taille des groupes au début d'Homo ne peut être déterminée avec certitude. Les groupes étaient probablement nombreux. Il est possible que les groupes soient au-dessus de la plage supérieure des tailles de groupe connues parmi les primates modernes. Les groupements contemporains de chimpanzés et de babouins comptent environ 100 individus ou plus. En 1993, les paléoanthropologues ont estimé que la taille des groupes d'Homo habilis et d'Homo rudolfensis aurait varié entre 70 et 85 individus. Les estimations étaient basées sur la taille du néocortex, car il existe une relation connue entre la taille du néocortex et la taille du groupe chez les primates non humains modernes. Avec le facteur supplémentaire de la bipédie, qui est énergétiquement moins coûteux que la quadripédie, la taille maximale du groupe écologiquement tolérable aurait pu être encore plus grande. En utilisant la même méthodologie, la taille estimée des groupes d'Homo ergaster était comprise entre 91 et 116 individus.
Le comportement social et de contre-attaque des espèces Homo antérieures s'est probablement transmis à Homo ergaster. Homo ergaster a probablement affiné et développé encore plus ces tactiques. Homo ergaster a probablement été la première espèce humaine à accéder au créneau des carnivores sociaux, ou chasseurs-cueilleurs. Un tel comportement aurait probablement été le résultat de contre-attaques dans un contexte de compétition alimentaire avec d'autres carnivores. Cela a probablement évolué à partir de quelque chose qui s’apparente à la chasse opportuniste parfois pratiquée par les chimpanzés. Le passage à la prédation en groupe aurait pu déclencher une cascade de changements évolutifs qui ont modifié le cours de l’évolution humaine.
Des comportements coopératifs tels que la chasse opportuniste en groupe, la défense contre les prédateurs et la recherche de charognards conflictuels auraient été essentiels à la survie. Cela se traduirait progressivement par une transition fondamentale en psychologie. Le comportement typique de « coopération compétitive » présenté par la plupart des primates a finalement été défavorisé par la sélection naturelle. Les tendances sociales prenant sa place, comme la chasse et d'autres activités, seraient devenues de véritables efforts de collaboration. Le comportement de contre-attaque est généralement manifesté chez les mâles des primates modernes. On pense donc que la chasse sociale chez les humains archaïques était également probablement une activité principalement masculine. Les femelles effectuaient probablement d'autres types de recherche de nourriture, récoltant des aliments qui ne nécessitaient pas de chasse (c'est-à-dire des fruits, des noix, des œufs, etc.).
La chasse étant une activité sociale, les individus partageaient probablement la viande entre eux. Cela aurait renforcé les liens à la fois entre les chasseurs eux-mêmes et entre les chasseurs et le reste du groupe Homo ergaster. Les femelles partageaient probablement également ce qu’elles avaient récolté avec le reste du groupe. Cette évolution aurait pu conduire au développement d’amitiés homme-femme vers des liens de couple monogames opportunistes. La sélection sexuelle des femelles favorisait probablement les mâles capables de chasser. Le comportement social émergent qui en résulte aurait été transmis et amplifié au fil des générations.
La seule preuve directe de la composition du groupe Homo ergaster provient d'une série de sites à l'extérieur d'Ileret au Kenya. Là, 97 empreintes laissées il y a environ 1,5 million d'années par un groupe d'au moins 20 individus ont été préservées. D'après la taille des empreintes, l'une des traces semble avoir été celle d'un groupe entièrement composé de mâles. Il s'agissait peut-être d'un groupe de travail spécialisé, tel qu'une patrouille frontalière ou un groupe de chasse ou de recherche de nourriture. Si cette évaluation est correcte, cela suggérerait en outre une division des responsabilités entre hommes et femmes.
Dans les sociétés modernes de chasseurs-cueilleurs qui ciblent de grosses proies, des groupes de mâles sont généralement envoyés pour abattre ces animaux à haut risque. En raison du faible taux de réussite et du risque élevé de chasser de grosses proies, les groupes de femelles ont tendance à se concentrer sur des sources de nourriture plus fiables. Les premiers Homo ergaster ont hérité de la culture Oldowan d'outils des australopithèques et des espèces Homo antérieures. Cependant, Homo ergaster a rapidement appris à frapper des éclats de pierre beaucoup plus gros que ses prédécesseurs et les espèces Homo contemporaines. Il y a 1,65 million d'années, Homo ergaster avait créé les artefacts en grande quantité et les premières haches à main qui marquent la culture acheuléenne. Il y a 1,6 à 1,4 millions d’années, la nouvelle industrie de l’outillage était largement implantée en Afrique de l’Est.
Les outils acheuléens diffèrent des outils oldowiens dans la mesure où leurs formes principales étaient clairement délibérées. Les outils Oldowan étaient probablement principalement utilisés comme marteaux pour casser les os et en extraire la moelle. La forme formée par le noyau ne semble pas avoir beaucoup d’importance. Alors que les haches à main de la culture acheuléenne démontrent une intention de produire des objets étroits et pointus. Ils étaient généralement produits en forme de larme, ovale ou triangulaire. Une fois en place, l'industrie acheuléenne est restée inchangée tout au long de l'existence d'Homo ergaster. De tels outils ont été produits pendant plus d’un million d’années jusqu’à il y a environ 250 000 ans. Plus tard, les outils n’étaient finalement pas très différents des outils produits il y a 1,65 millions d’années.
Les assemblages acheuléens les plus anciens préservent également des formes de noyau similaires à celles des outils Oldowan. Cependant, il n’existe pas de véritables formes intermédiaires connues entre les deux. Cela suggère que l’apparition des outils acheuléens a été un développement brutal et soudain. Le développement le plus important qui a conduit aux outils acheuléens a probablement été l’apprentissage par les premiers hominidés de la capacité de frapper de gros éclats. Ces gros flocons mesuraient jusqu'à un pied ou plus de longueur. Ils provenaient de rochers plus gros. À partir de ces éclats plus gros, Homo ergaster pouvait fabriquer de nouveaux outils tels que des haches à main. Bien sûr, « hache à main » implique que toutes les haches à main étaient utilisées pour couper et étaient tenues à la main. Cependant, ils se présentaient sous différentes formes et tailles et remplissaient probablement plusieurs fonctions différentes.
Des haches à main soigneusement formées et symétriques peuvent avoir été lancées sur des proies semblables aux disques modernes. Des haches à main produites de manière plus occasionnelle peuvent simplement avoir servi de sources portables pour des éclats tranchants. Certains auraient pu être utilisés pour gratter ou couper du bois. De plus, les haches à main étaient des outils de boucherie efficaces. Ils étaient peut-être également utilisés pour démembrer les carcasses de gros animaux. Il existe des haches à main préservées qui sont trop lourdes et trop grandes pour être utilisées à des fins pratiques apparentes. On a également découvert des collections de centaines de haches à main sans signes évidents d'utilisation.
L'utilisation des haches à main plus grandes ainsi que des collections de haches à main apparemment inutilisées est spéculative et conjecturale. Une idée populaire dans les médias et même fréquemment citée dans le monde universitaire est que de grandes et impressionnantes haches à main auraient pu être des emblèmes utilisés pour attirer des partenaires. Il est suggéré que les fabricants de ces grands axes fassent ainsi preuve de force, de coordination et de détermination. Il est postulé que ces attributs dérivés de la production de grandes haches à main auraient pu être considérés comme attrayants.
En 2009, les paléoanthropologues ont noté que même si cette théorie était « à la fois intrigante et émotionnellement attrayante », il y avait peu de preuves à l'appui et la théorie était invérifiable. Ils considéraient qu'il était plus probable que les variations de la morphologie des haches au cours de centaines de milliers d'années soient le résultat d'une myriade de facteurs différents. Les variations et les haches « géantes » étaient plus probablement le résultat d’essais et d’erreurs plutôt que d’un facteur unique et primordial dans la sélection sexuelle. À mesure qu'Homo a migré vers des environnements de savane ouverte, les rencontres avec des incendies naturels ont dû devenir plus fréquentes et plus importantes.
Il est possible qu’Homo ergaster ait été le premier humain à maîtriser le contrôle du feu. Il est également possible, voire probable, qu'ils aient utilisé le feu pour cuisiner. La cuisson rend les aliments carnés et végétaux plus digestes. Cela aurait très bien pu être important puisque les intestins d’Homo ergaster étaient de taille réduite par rapport à ceux de leurs ancêtres. On suppose souvent que Homo ergaster et/ou Homo erectus ont été les premiers Homo à contrôler le feu. Cependant, les preuves solides manquent quelque peu dans les archives fossiles. Cela est peut-être dû en partie à la difficulté ou à l’improbabilité de conserver des preuves réelles de l’utilisation du feu.
Deux des premiers sites généralement considérés comme préservant des preuves de l'utilisation du feu sont deux sites situés près du lac Baringo, au Kenya. Les deux sites sont datés de 1,5 million d’années. Sur un site, les preuves consistent en des sédiments brûlés et des outils en pierre altérés par la chaleur. Sur le deuxième site, de gros fragments d'argile cuite sont conservés, associés à des outils en pierre et à des restes fauniques. Bien qu'il soit difficile d'exclure une origine naturelle des résidus d'incendie mis en évidence, les sites restent de bons candidats pour une utilisation précoce en cas d'incendie.
Plusieurs sites préservant des preuves plus largement acceptées de l'utilisation du feu ont été datés d'il y a 1 million d'années ou moins. Celles-ci sont datées d'après l'émergence et la dernière mention généralement acceptée d'Homo ergaster. Il s'agit notamment de sites grottes tels que Wonderwerk et Swartkrans en Afrique du Sud, et de sites ouverts, tels que Kalambo Falls en Zambie. Le site Gesher Benot Ya'aqov en Israël est daté d'il y a environ 700 000 ans. Il préserve des preuves largement acceptées de l’utilisation du feu à travers des matériaux brûlés et des microartefacts en silex brûlé. Ces preuves sont préservées à de nombreux niveaux sédimentaires. Depuis environ 400 000 ans, les traces d'incendies deviennent encore plus nombreuses sur des sites en Afrique, en Europe et en Asie.
La moelle épinière de Turkana Boy aurait été plus étroite que celle des humains modernes. Cela suggère que le système nerveux d'Homo ergaster, y compris ses muscles respiratoires, n'a peut-être pas été avancé au degré nécessaire pour produire ou contrôler la parole. Cependant, en 2001, des anthropologues ont suggéré que Turkana Boy souffrait de dysplasie squelettique et de scoliose. Si cela était vrai, il s’ensuit qu’il n’aurait pas été représentatif du reste de son espèce. En 2006, un anthropologue a décrit un spécimen d'Homo erectus provenant de Dmanisi, en Géorgie. Le spécimen d'Homo erectus était daté de 1,78 millions d'années.
Le fossile préserve les plus anciennes vertèbres Homo connues et la colonne vertébrale trouvée. Les restes fossilisés relèvent de la portée de la colonne vertébrale humaine moderne. Cela suggère que l’individu aurait été capable de parler. Meyer et ses collègues ont conclu que la parole était probablement possible au sein d'Homo très tôt. Leurs conclusions confirment que Turkana Boy souffrait probablement d'une anomalie congénitale, peut-être d'une sténose vertébrale. Cependant, en 2013 et 2014, une autre équipe d'anthropologues a conclu qu'il n'y avait aucune preuve d'anomalies congénitales chez Turkana Boy. Contrairement aux études de 2001 et 2006, l'équipe a considéré le spécimen comme représentatif de l'espèce [Wikipedia].
: Nous expédions toujours des livres au niveau national (aux États-Unis) via USPS ASSURÉ courrier des médias (« tarif livre »). Il existe également un programme de réduction qui peut réduire les frais de port de 50 à 75 % si vous achetez environ une demi-douzaine de livres ou plus (5 kilos et plus). Nos frais de port sont aussi raisonnables que les tarifs USPS le permettent. ACHATS SUPPLÉMENTAIRES recevez un TRÈS GRAND
Votre achat sera généralement expédié dans les 48 heures suivant le paiement. Nous emballons aussi bien que n'importe qui dans le secteur, avec de nombreux rembourrages et conteneurs de protection. Le suivi international est fourni gratuitement par l'USPS pour certains pays, d'autres pays entraînent des frais supplémentaires.
Nous proposons le courrier prioritaire du service postal américain, le courrier recommandé et le courrier express pour les envois internationaux et nationaux, ainsi que United Parcel Service (UPS) et Federal Express (Fed-Ex). Merci de demander un devis. Veuillez noter que pour les acheteurs internationaux, nous ferons tout notre possible pour minimiser votre responsabilité en matière de TVA et/ou de droits. Mais nous ne pouvons assumer aucune responsabilité pour les taxes ou droits qui pourraient être perçus sur votre achat par le pays de votre résidence. Si vous n'aimez pas les systèmes de taxes et de droits imposés par votre gouvernement, veuillez vous plaindre auprès d'eux. Nous n'avons aucune capacité à influencer ou à modérer les systèmes fiscaux/droits de votre pays.
Si à la réception de l'article vous êtes déçu pour quelque raison que ce soit, je propose une politique de retour de 30 jours sans poser de questions. Nous n'avons évidemment aucune possibilité d'influencer, de modifier ou de renoncer aux politiques d'eBay.
À PROPOS: Avant notre retraite, nous voyageions plusieurs fois par an en Europe de l'Est et en Asie centrale à la recherche de pierres précieuses et de bijoux anciens provenant des centres de production et de taille de pierres précieuses les plus prolifiques du monde. La plupart des articles que nous proposons proviennent d'acquisitions que nous avons réalisées au cours de ces années en Europe de l'Est, en Inde et au Levant (Méditerranée orientale/Proche-Orient) auprès de diverses institutions et revendeurs. Une grande partie de ce que nous générons sur Etsy, Amazon et Ebay est destinée à soutenir des institutions dignes d'Europe et d'Asie liées à l'anthropologie et à l'archéologie. Bien que nous ayons une collection de pièces de monnaie anciennes se comptant par dizaines de milliers, nos principaux intérêts sont les bijoux anciens et les pierres précieuses, reflet de notre formation universitaire.
Bien qu’elles soient peut-être difficiles à trouver aux États-Unis, en Europe de l’Est et en Asie centrale, les pierres précieuses antiques sont généralement démontées des anciennes montures cassées – l’or est réutilisé – les pierres précieuses sont recoupées et réinitialisées. Avant que ces magnifiques pierres précieuses antiques ne soient retaillées, nous essayons d’acquérir les meilleures d’entre elles dans leur état d’origine, antique et fini à la main – la plupart d’entre elles ont été fabriquées il y a un siècle ou plus. Nous pensons que le travail créé par ces maîtres artisans disparus depuis longtemps mérite d'être protégé et préservé plutôt que de détruire ce patrimoine de pierres précieuses antiques en découpant l'œuvre originale de l'existence. En préservant leur travail, d’une certaine manière, nous préservons leur vie et l’héritage qu’ils ont laissé aux temps modernes. Il vaut bien mieux apprécier leur métier que de le détruire avec une coupe moderne.
Tout le monde n’est pas d’accord : au moins 95 % des pierres précieuses antiques qui arrivent sur ces marchés sont retaillées et l’héritage du passé est perdu. Mais si vous êtes d'accord avec nous que le passé mérite d'être protégé et que les vies passées et les produits de ces vies comptent toujours aujourd'hui, envisagez d'acheter une pierre précieuse naturelle antique, taillée à la main plutôt qu'une pierre précieuse taillée à la machine produite en série (souvent synthétique). ou « produites en laboratoire ») qui dominent le marché aujourd’hui. Nous pouvons sertir la plupart des pierres précieuses antiques que vous achetez chez nous dans votre choix de styles et de métaux allant des bagues aux pendentifs en passant par les boucles d'oreilles et les bracelets ; en argent sterling, en or massif 14 carats et en or 14 carats. Nous serions heureux de vous fournir un certificat/garantie d’authenticité pour tout article que vous achetez chez nous. Je répondrai toujours à chaque demande, que ce soit par e-mail ou par message eBay, alors n'hésitez pas à écrire.
| Publisher | Time-Life Books (1972) |
| Dimensions | 10¾ x 8¾ inches; 1¾ pounds |
| Format | Hardcover w/printed, decorated laminate covers |
| Color | Multi-Color |
| Material | Paper |
| Length | 160 pages |
| Original/Reproduction | Original |